- y Library
- Complete Notes ..
- जीव विज्ञान (Bi ..
- पादप कार्यिकी
- पादप कार्यिकी
पादप कार्यिकी
परासरण (Osmosis) या परासरणीय विसरण (Osmotic Diffusion)–
– जब दो विभिन्न सान्द्रता वाले विलयनों को अर्द्ध-पारगम्य झिल्ली के द्वारा पृथक् कर दिया जाता है तब विलायक का कम सान्द्रता वाले विलयन से अधिक सान्द्रता वाले विलयन की तरफ अर्द्ध-पारगम्य झिल्ली से होकर जाना, परासरण (Osmosis) कहलाता है।
– अर्द्ध-पारगम्य झिल्ली के द्वारा (H2O) विलायक का अपनी अधिक सान्द्रता से कम सान्द्रता की ओर जाना परासरण कहलाता है।
– परासरण के अंतर्गत जब कोशिका में जल का प्रवेश होता है तो इसे अन्त:परासरण (Endosmosis) कहते हैं (किशमिश का पानी में फूलना) तथा जब कोशिका से जल बाहर निकलता है तो इसे परासरण (Exomosis) कहते हैं। अंगूरों को (Hyperthonic) घोल में रखना।

परासरण दाब (Osmotic Pressure or O.P.) –
– परासरण दाब (O.P.) विलयन में उत्पन्न वह दाब या खिंचाव है जो एक अर्द्ध-पारगम्य झिल्ली द्वारा विलयन एवं जल को पृथक् करने पर विलयन में विकसित होता है।
– शुद्ध जल का परासरण दाब (O.P.) शून्य होता है। यह घोल में विलेय का गुण है।
– घोल में विलेय (Solute) पदार्थों की सान्द्रता जितनी अधिक होगी घोल का परासरण दाब भी उतना ही अधिक होगा। अत: इसका मान धनात्मक होता है।
परासरण का महत्त्व (Significance of Osmosis) –
– जड़ों में मूलरोम मृदा से परासरण की क्रिया द्वारा जल का अवशोषण करते हैं।
– पौधों में एक कोशिका से दूसरी कोशिका में जल का चालन तथा पौधों में जल का वितरण परासरण द्वारा होता है।
– परासरण की क्रिया के फलस्वरूप उत्पन्न स्फीति (Turgidity) पत्तियों, तने, पुष्पों में निश्चित आकृति बनाए रखने में सहायता करती है तथा पौधों को यांत्रिक शक्ति प्रदान करती है।
– रन्ध्रों का खुलना तथा बन्द होना परासरण की क्रिया पर निर्भर करता है। (अंत: परासरण व बाह्य परासरण)
पादप जल अवशोषण (Water Absorption) –
– जल जीवद्रव्य का एक अत्यन्त आवश्यक भाग होता है। प्रोटोप्लाज्म के सम्पूर्ण भार का 80 से 90% भाग जल होता है। जल एक श्रेष्ठ विलायक है तथा यह पादपों के विभिन्न कार्यिकी प्रक्रियाओं में व खनिज पोषकों के ग्रहण व वितरण में प्रमुख भूमिका निभाता है। सभी कार्बनिक एवं अकार्बनिक पदार्थों का अभिगमन जल द्वारा ही होता है।
जल के रूप (Forms of Water) –
– भूमि को जल मुख्य रूप से वर्षा से प्राप्त होता है। कुछ जल तो बहकर जलाशयों में चला जाता है। इसे अपवाहित जल (Run off Water) कहते हैं। शेष जल रिस-रिस कर (Percolate Water) भूमि में चला जाता है।
(a) गुरुत्वीय जल (Gravitational Water) – गुरुत्वाकर्षण के कारण यह जल गहराई में चला जाता है। सामान्य रूप से पादपों को उपलब्ध नहीं होता। कुछ Xerophytes जैसे – Calotropis, Prosopis, Capparis अवशोषित कर सकते हैं।
(b) आर्द्रताग्राही जल (Hygroscopic Water) – मिट्टी के कणों की सतह पर जल की सूक्ष्म अधिशोषित (Adsorbed) परत को आर्द्रता जल कहते हैं। यह जल भी पौधों को उपलब्ध नहीं होता है। इस जल का अत्यधिक –ve होता है।
(c) रासायनिक बंधित जल (Chemically Combined Water) – मृदा के कणों में विद्यमान रासायनिक यौगिकों में रहने वाली जल की मात्रा ही रासायनिक जल होता है। यह भी पौधों को उपलब्ध नहीं होता है।
(d) कोशिका जल (Capillary Water) – मिट्टी के कणों के बीच बनी हुई कोशिकाओं में जल की मात्रा को कोशिका जल कहते हैं। यही जल पादपों की जड़ों के लिए उपयोगी होता है अर्थात् यह जल पौधों को उपलब्ध होता है। पादप इसी जल को अवशोषित करते हैं।
– होलार्ड (Holard) – यह जल की वह सम्पूर्ण मात्रा है जो मृदा में स्थित होती है।
होलार्ड = क्रेसार्ड + इकार्ड (Holard = Chresard + Echard)
– क्रेसार्ड (Chresard) – मृदा में पाए जाने वाला वह जल जो पादपों को उपलब्ध होता है।
– इकार्ड (Echard) – यह वह जल है जो कि पौधों द्वारा अवशोषित नहीं होता है।
जल अवशोषण अंग (Organ for Water Absorption)–
– निम्न श्रेणी के पादप जल को सम्पूर्ण सतह या मूलाभासों (Rhizoids) से अवशोषित करते हैं परन्तु टेरिडोफाइट्स व स्पर्मेटोफाइट्स (बीज धारी पादप) में जल अवशोषण मूल (Root) द्वारा होता है।
जल अवशोषण का मार्ग (Path of Water Absorption)–
– भूमि में उपस्थित जल को जड़ों के जाइलम तक पहुँचना होता है। भूमि में जल के संपर्क में मूलरोम होते हैं। मूलरोम की पैक्टिन सतह पर जल सर्वप्रथम अधिशोषित होता है।
– तत्पश्चात् जल मूलरोम की एपिडर्मिस में आता है यहाँ से यह वल्कुट में होता हुआ वल्कुट की आन्तरिक तह अन्त:श्चर्म या एण्डोडर्मिस में पहुँचता है। अन्तस्त्वचा (Endodermis) की भित्तियाँ सुबेरीनीकृत (केस्पेरीयन पट्टियाँ) होती है परन्तु जल को जाइलम तक पहुँचाने के लिए प्रोटोजाइलम के विपरीत पतली भित्ति वाली ‘पथ कोशिकाएँ (Passage Cell)’ होती है। यहाँ से जल पतली भित्ति युक्त परिरम्भ कोशिकाओं में से होता हुआ जाइलम तक पहुँच जाता है।
Soil Solution à Root hair (Epiblema) à Cortex à Endodermis (Passage Cell) à Pericycle à Protoxylem à Metaxylem.
– सिम्प्लास्ट या संवलक पथ (Symplast) – सतत जीवित अंश से बने मार्ग को सिम्प्लास्ट कहते हैं। यह जीवित मार्ग होता है। जल कोशिका झिल्ली से कोशिका द्रव्य (Cytoplasm) में पहुँचाता है। कोशिका द्रव्य व जीवद्रव्य तन्तु मिलाकर यह अंश (Symplast) बनाते हैं। कोशिका के भीतर होकर प्लाज्मोडेस्मोटा से जल का परिवहन सिम्प्लास्ट पाथ कहलाता है। सिम्प्लास्ट पाथ के दौरान जल कर परिवहन झिल्लियों द्वारा भी होता है। जिसे पारकला पथ (Transmembrane Path) भी कहा जाता है।
– एपोप्लास्ट या अपसंवलक पथ (Apoplast) – यह पादप का अजीवित भाग होता है। कोशिका भित्ति तथा अन्तरा कोशिकीय अवकाश तथा जाइलम गुहिका मिलकर ‘एपोप्लास्ट’ का निर्माण करते हैं। ये शब्द मुंच ने दिए थे।
– मूलरोम से Cortex तक जल का प्रवाह Apoplastic या Symplastic हो सकता है। Endodermis में Casparian Strips होती है। अत: जल का गमन Symplast से ही होता है।
जल अवशोषण को प्रभावित करने वाले कारक –
प्राप्य मृदा जल (Available Soil Water) –
– पादप भूमि में उपस्थित कोशिका जल (Capillary Water) को अवशोषित करते हैं। जल अवशोषण मृदा में उपस्थित कोशिका जल की मात्रा बढ़ने पर बढ़ जाता है।
– अगर मृदा से अधिक जल उपस्थित है, तो ऐसी मृदा को जलाक्रांत (Water Logged Soil) मृदा कहते हैं। यह मृदा कार्यिकी रूप से शुष्क (Physiologically Dry) होती है। इसमें वायु अनुपस्थित होती है। इस कारण ये ऐसी मृदा में जड़ों में अवायवीय श्वसन होता है व एल्कोहॉल का निर्माण होता है। एल्कोहॉल बनने के कारण जड़े नष्ट हो जाती हैं। जल मग्न खेतों में एल्कोहॉल की गंध आती है। (मरुस्थलीय मृदा भौतिक शुष्क होती है।)
मृदा तापमान (Soil Temperature) –
– मृदा तापमान निम्नलिखित क्रियाओं को प्रभावित करता है। (अनुकूल ताप 20-35oC)
(i) यह कोशिका झिल्ली की पारगम्यता को प्रभावित करता है। कम ताप से कोशिका झिल्ली की पारगम्यता घट जाती है।
(ii) यह एन्जाइमों की सक्रियता के लिए आवश्यक होता है। तभी मूलरोम निर्माण व जड़ों की वृद्धि होगी।
(iii) मृदा तापमान कोशिका जल की श्यानता (Viscosity) को प्रभावित करता है। कम ताप से जल की श्यानता बढ़ती है।
मृदा वायु (Soil Air) –
– अच्छे वातायन वाली मृदा में जल अवशोषण अधिक होता है।
– अल्प वातित मृदा में जड़ों के श्वसन हेतु पर्याप्त ऑक्सीजन नहीं होता है। अत: अल्पवातित मृदा को कार्यिकी शुष्क कहा जाता है।
मृदा के लवण (Soil Salt) –
– जल अवशोषण की दर मृदा में उपस्थित लवणों की सान्द्रता के व्युत्क्रमानुपाती होती है। लवणीय मृदा कार्यिकी शुष्क होगी।
– उपयुक्त मृदा घोल में ही जल अवशोषण होता है। मृदा में लवणों की सान्द्रता बढ़ने पर जल अवशोषण की दर घट जाती है। इसी कारण लवणीय भूमि (Saline Soil) कार्यिकी दृष्टि से शुष्क (Physiologically Dry) होती है। इस मृदा में केवल लवणोद्भिद (Halophytes) पादप ही उगते हैं, क्योंकि उनका O.P. अधिक बनाए रखते हैं।
वाष्पोत्सर्जन (Transpiration) –
– क्रेमर (Kramer) के अनुसार जल अवशोषण की दर सीधे ही वाष्पोत्सर्जन के समानुपाती होती है।
– वाष्पोत्सर्जन की दर बढ़ने पर जल अवशोषण की क्रिया बढ़ जाती है, क्योंकि वाष्पोत्सर्जन के कारण निष्क्रिय जल अवशोषण बढ़ जाता है।
जल अवशोषण की कुछ अन्य विधियाँ –
– माइकोराइजा (Mycorrhiza) द्वारा – कुछ शंकुनुमा वृक्षों (Conifers) में मूलरोम विकसित नहीं होते हैं। अत: मूलरोमों के स्थान पर जड़ों में कवक सूत्रों के साथ संबंध स्थापित हो जाता है जिसे माइकोराइजा कहते हैं। यह सहजीवन का उदाहरण होता है। जिसमें कवक सूत्र जड़ों के अवशोषक अंग होते हैं। ये कवक सूत्र जल तथा खनिज लवणों का अवशोषण करके पौधों की जड़ों को देते हैं।
– वेलामेन (Velamen) द्वारा – यह एपीफाइट्स (Epiphytes) या उपरिरोही पादपों में जैसे – आर्किड (Orchid) तथा ऐस्पेरेगस (Asparagus) में पाया जाता है।
– इन पादपों में वायुमण्डल की जल वाष्प को, लटकने वाली जड़ों (Hanging Roots) द्वारा अवशोषित किया जाता है। इन जड़ों में वल्कुट के बाहर वेलामेन नामक ऊत्तक स्थित होता है।
रसारोहण (Ascent of Sap) –
– पौधे के भूमिगत भाग (जड़) से अवशोषित जल के गुरुत्वाकर्षण के विपरीत पौधे के वायुवीय भागों तक पहुँचने की क्रिया को रसारोहण कहते हैं।
मूल दाब वाद (Root Pressure Theory) –
– प्रिस्टले द्वारा दिया गया था।
– इस सिद्धांत के अनुसार मूल दाब जाइलम रस में उत्पन्न होने वाला वह दाब है जो कि मूल वल्कुट की स्फीति कोशिकाओं द्वारा उत्पन्न होता है। यह धनात्मक दाब है जो Xylem रस को ऊपर की ओर धकेलता है।
वाष्पोत्सर्जनकर्षणा–संसजन बल सिद्धांत (Transpiration Pull – Cohesive Force Theory) –
– डिक्सन व जोली द्वारा प्रस्तुत की गई।
– यह रसारोहण का सर्वाधिक व सर्वमान्य सिद्धांत है।
– इस सिद्धांत के अनुसार रसारोहण की क्रिया में तीन घटक काम आते हैं।
(i) ससंजन – जल के अणुओं के मध्य एक विशेष आकर्षण बल होता है जिस ससंजन बल (Cohesion) कहते हैं। यह बल जाइलम में एक निरंतर जल स्तम्भ का निर्माण करता है।
(ii) आसंजन – जल के अणुओं व जाइलम भित्ति के मध्य एक बल कार्य करता है। जिसे आसंजन बल (Adhesion) कहते हैं, जो कि जाइलम में उपस्थित निरंतर जल स्तम्भ का रख-रखाव करता है।
(iii) वाष्पोत्सर्जन कर्षण – पत्तियों में तीव्र वाष्पोत्सर्जन DPD की अधिकता के कारण से जाइलम में एक –ve दबाव उत्पन्न होता है जिसे वाष्पोत्सर्जन कर्षण (Transpiration Pull) कहते हैं और यह जाइलम के जल स्तम्भ में पानी को ऊपर खींचने के लिए उत्तरदायी होता है। अत: रसारोहण ससंजन, आसंजन व वाष्पोत्सर्जन कर्षण तीनों का ही सम्मिलित प्रभाव है।
वाष्पोत्सर्जन (Transpiration)
– सजीव पौधों के वायवीय (पुष्प, फल, पत्ति, तना) भागों से जल के वाष्प के रूप में त्याग को वाष्पोत्सर्जन (Transpiration) कहते हैं।
– पादपों में सबसे कम वाष्पोत्सर्जन मासलोद्भिद मरुद्भिदों (Succulent Xerophyte) में पाया जाता है।
– सबसे अधिक वाष्पोत्सर्जन मीसोफाइट्स में (Mesophytes) पाया जाता है।
वाष्पोत्सर्जन के प्रकार (Type of Transpiration) –
– वाष्पोत्सर्जन निम्नलिखित तीन प्रकार का होता है-
(i) रन्ध्रीय वाष्पोत्सर्जन (Stomatal Transpiration) – पौधों की पत्तियों तथा अन्य कोमल अंगों पर उपस्थित रन्ध्रों से होने वाले वाष्पोत्सर्जन को रन्ध्रीय वाष्पोत्सर्जन कहते हैं। यह सबसे अधिक होता है। कुल वाष्पोत्सर्जन का 80 से 90% वाष्पोत्सर्जन रन्ध्रों द्वारा होता है।
(ii) उपत्वचीय वाष्पोत्सर्जन (Cuticular Transpiration) – यह वाष्पोत्सर्जन शाकीय तनों (Herbaceous) तथा पत्तियों पर पाए जाने वाली उपत्वचा द्वारा होता है क्योंकि इनकी Epidermis पर पतली मोम जैसी परत Cuticle पाई जाती है। उपत्वचीय वाष्पोत्सर्जन लगभग 9% होता है।
(iii) वातरुध्रीय वाष्पोत्सर्जन (Lenticular Transpiration) – काष्ठीय पौधों के तनों, छाल, कुछ फलों की त्वचा में वातरन्ध्र पाए जाते हैं। वातरन्ध्रों द्वारा भी अल्प मात्रा में जलवाष्प के रूप में जल की हानि होती है। इसे वातरन्ध्रीय वाष्पोत्सर्जन कहते हैं।
– पत्रक वाष्पोत्सर्जन (Foliar Transpiration) – पत्तियों से होने वाला सम्पूर्ण वाष्पोत्सर्जन पत्रक वाष्पोत्सर्जन कहलाता है। (Foliar Transpiration) – रंध्रीय वाष्पोत्सर्जन + उपत्वचीय वाष्पोत्सर्जन, पत्तियों से।
– शैवाल (Algae) कवक (Fungi) व निमग्न जलीय पादपों (Submerged plant) में वाष्पोत्सर्जन नहीं होता है।
– शाकीय पौधों के कोमल वायुवीय अंगों तथा पत्तियों की बाह्य त्वचा पर छिद्र के रूप में रन्ध्र पाए जाते हैं। यह रन्ध्री छिद्र दो विशेष प्रकार की बाह्य त्वचीय कोशिकाओं (Epidermal Cells) से घिरा होता है, जिन्हें द्वार कोशिकाएँ या रक्षक कोशिका (Guard Cell) कहते हैं। इनकी आकृति वृक्काकार (Kidney Shaped) होती है। एक पर्णरन्ध्र में द्वार कोशिकाओं या रक्षक कोशिकाओं की संख्या दो होती है। एक बीज पत्रियों में ये डम्बलाकार होती है। द्वार कोशिकाओं में हरित लवक होते हैं।
– ग्रेमिनी (Gramineae) या घास कुल के पादपों में रक्षक कोशिकाओं की आकृति ‘डम्बलाकार’ (Dumbbell Shaped) होती है।
– प्रत्येक द्वार कोशिकाओं की बाह्य भित्ति (Outer) पतली (Thin) तथा प्रत्यास्थ (Elastic) होती है जबकि आन्तरिक भित्ति (Inner Wall) मोटी (Thick) तथा अप्रत्यास्थ (Non-Elastic) होती है। अधिक Cellulose जमने से।
– रन्ध्रों को चारों ओर से कुछ विशिष्ट बाह्य त्वचीय कोशिकाएँ घेरे रहती हैं, जिन्हें सहायक कोशिकाएँ (Subsidiary Cells) या अतिरिक्त (Accessory) कोशिकाएँ कहते हैं।
– रन्ध्र वायु कोष्ठों से जुड़े रहते हैं तथा इनसे बनी गुहा को अधोरन्ध्री गुहा (Sub Stomatal Cavity) कहते हैं।
– यह सिद्धांत Levitt (1973-74) द्वारा दिया गया, जो कि रंध्रीय का आधुनिक व मान्यता प्राप्त सिद्धांत है।
सर्वप्रथम Funjino ने बताया कि रन्ध्र खुलते समय रक्षक कोशिकाओं (Guard Cells) में K+ आयनों का अन्तर्वाह होता है।

– रन्ध्रगति की क्रियाविधि (Mechanism of Stomatal Movement) –

रंध्र का खुलना तथा बंद होना
– रन्ध्रों के खुलने व बन्द होने के लिए रक्षक कोशिकाओं की स्फीति व स्थिलता (rigidity & flacidity) उत्तरदायी है, जो इन कोशिकाओं के परासरणी दाब पर निर्भर करती है। रक्षक कोशिकाओं का परासरणी दाब कई कारकों; जैसे – शर्करा सान्द्रता, CO2 सान्द्रता, आयनों की सान्द्रता, pH का मान आदि पर निर्भर करता है। इस सम्बन्ध में वैज्ञानिकों द्वारा समय-समय पर प्रस्तुत विभिन्न मत निम्नलिखित प्रकार हैं –
प्रकाश संश्लेषी सिद्धान्त (Photosynthetic theory) –
– वॉन मोल (Von Mohl, 1856) के अनुसार प्रकाश में रक्षक कोशिकाओं के क्लोरोप्लास्ट प्रकाश संश्लेषण द्वारा परासरणी सक्रिय पदार्थों का निर्माण करते हैं, जिसके फलस्वरूप इनका परासरणी दाब बढ़ जाता है। ये कोशिकाएँ जल अवशोषित कर स्फीति (turgid) हो जाती हैं, जिससे रन्ध्र खुल जाते हैं। रात्रि में प्रकाश संश्लेषण के अभाव में रक्षक कोशिकाओं का परासरणी दाब कम होने से शिथिल (flacid) हो जाती है तथा रन्ध्र बन्द हो जाते हैं।
दोष (Objections) –
– इस मत के अनुसार पत्ती के वातावरण में प्रकाश की उपस्थिति में CO2 सान्द्रता बढ़ाने पर अधिक रन्ध्र खुलने चाहिए जबकि रन्ध्र आंशिक रूप से बन्द हो जाते हैं।
– अधिकतर पौधों में रक्षक कोशिकाएँ प्रकाश संश्लेषण क्रिया द्वारा पर्याप्त शर्करा एकत्र नहीं कर पाती हैं।
– कुछ पौधों में रन्ध्र रात्रि में खुले रहते हैं जबकि दिन में जब प्रकाश संश्लेषण होता है, रन्ध्र बन्द रहते हैं।
स्टार्च-शर्करा अन्त:परिवर्तन मत
(Starch-sugar interconversion theory) –
– लॉयड (Lloyed, 1908) के अनुसार रक्षक कोशिकाओं में स्टार्च की मात्रा दिन में कम तथा रात में अधिक होती है। स्टार्च तथा मण्ड के अंत:परिवर्तन से रक्षक कोशिकाओं की परासरण सान्द्रता (osmotic concentration) प्रभावित होती है। प्रकाश की उपस्थिति में स्टार्च का शर्करा में परिवर्तन होता है, जिससे रक्षक कोशिकाओं की परासरण सान्द्रता बढ़ जाती है। रक्षक कोशिकाएँ जल अवशोषित कर स्फीति हो जाती हैं तथा रन्ध्र खुल जाते हैं। इसके विपरीत अंधकार में शर्करा का परिवर्तन स्टार्च में हो जाता है, जिससे इनकी परासरणी सान्द्रता कम हो जाती है। इस अवस्था में रक्षक कोशिकाएँ शिथिल हो जाती हैं तथा रन्ध्र बन्द हो जाते हैं।
– सायरे (Sayre, 1926) के अनुसार स्टार्च-शर्करा का अन्त:परिवर्तन pH पर निर्भर करता है। प्रकाश में (दिन में) श्वसन क्रिया द्वारा मुक्त CO2 पत्ती की पर्णमध्योत्तक कोशिकाओं द्वारा प्रकाश संश्लेषण में उपभोग कर ली जाती है, जिससे उपरन्ध्रीय गुहा में CO2 सान्द्रता कम हो जाती है। इसके फलस्वरूप रक्षक कोशिकाओं में pH का मान बढ़ जाता है (pH 7-7.5)। इस क्षारीय स्थिति (basic condition) में स्टार्च का अपघटन शर्करा में हो जाता है। शर्करा के कारण रक्षक कोशिकाओं का परासरणी दाब बढ़ जाता है जिससे ये जल अवशोषण कर स्फित हो जाती है तथा रन्ध्र खुल जाते हैं । रात्रि में (अंधकार में) प्रकाश संयोजन के अभाव में श्वसन में मुक्त CO2 का उपभोग नहीं हो पाता है तथा यह अन्तर्कोशीय अवकाशों में एकत्र हो जाती है। CO2 की अधिक सान्द्रता के कारण रक्षक कोशिकाओं में pH का मान कम (4-4.5) हो जाता है। इस अम्लीय स्थिति (acidic condition) में शर्करा का स्टार्च में परिवर्तन हो जाता है, जिससे रक्षक कोशिकाओं का परासरणी दाब कम हो जाता है जिससे ये जल त्याग कर स्थिल हो जाती हैं तथा रन्ध्र बन्द हो जाते हैं।
– प्रकाश व अन्धकार में द्वार कोशिकाओं (रक्षक कोशिकाओं) में होने वाले परिवर्तनों को यहाँ सारणीबद्ध किया गया है–
| प्रकाश में (Light) | अन्धकार में (Dark) |
| 1. श्वसन में मुक्त अन्तर्कोशिक अवकाशों में स्थित CO2 का प्रकाश संश्लेषण में उपभोग। | श्वसन द्वारा मुक्त CO2 का प्रकाश संश्लेषण के अभाव में अन्तर्कोशिक अवकाशों में एकत्र होना। |
| 2. रक्षक कोशिकाओं में pH का मान (7) बढ़ता है (क्षारीय स्थिति उत्पन्न होना)। | रक्षक कोशिकाओं में pH का मान (4.5) घटता है (अम्लीय स्थिति उत्पन्न होना)। |
| 3. pH बढ़ने से स्टार्च का फॉस्फोराइलेज द्वारा शर्करा में अपघटन। | pH घटने से शर्करा का स्टार्च में परिवर्तन होता है। |
| 4. शर्करा के कारण रक्षक कोशिकाओं का परासरणी दाब बढ़ना। | स्टार्च के परासरणी निष्क्रिय होने के कारण रक्षक कोशिकाओं का परासरणी दाब कम हो जाता है। |
| 5. पास की कोशिकाओं से जल का अन्तःपरासरण द्वारा रक्षक कोशिकाओं में प्रवेश। | रक्षक कोशिकाओं से जल बहिःपरासरण द्वारा पास की कोशिकाओं में प्रवेश कर जाता है। |
| 6. रक्षक कोशिकाओं के स्फीति होने से रन्ध्र का खुलना। | रक्षक कोशिकाएँ शिथिल हो जाती हैं तथा रन्ध्र बन्द हो जाते हैं। |
– स्टीवर्ड (Steward, 1964) ने इस परिकल्पना को परिवर्तित किया। स्टीवर्ड के अनुसार ग्लूकोज-1-फॉस्फेट रक्षक कोशिकाओं के परासरणी दाब को परिवर्तित करने में असमर्थ होता है। रक्षक कोशिकाओं का परासरणी दाब तब ही बढ़ता है, जब ग्लूकोज-1-फॉस्फेट का ग्लूकोज तथा अकार्बनिक फॉस्फेट में परिवर्तन होता है। स्टीवर्ड के अनुसार ग्लूकोज-1-फॉस्फेट का रूपान्तरण ग्लूकोज-6-फॉस्फेट में होता है तथा ग्लूकोज-6-फॉस्फेट का फॉस्फेटेज नामक एन्जाइम द्वारा ग्लूकोज व अकार्बनिक फॉस्फेट में परिवर्तन होता है।

– इस प्रकार रन्ध्र के बन्द होते समय ATP की आवश्यकता होती है, जो सम्भवतः श्वसन द्वारा उपलब्ध होती है, इसलिए यह एक सक्रिय क्रिया है।

रन्ध्र खुलने व बन्द होने की स्टीवर्ड परिकल्पना का आरेखी निरूपण
– दोष (Objections) – स्टार्च-शर्करा परिकल्पना निम्नलिखित कारणों से सर्वमान्य नहीं है –
– कुछ पौधों की रक्षक कोशिकाओं में (एकबीजपत्रियों में) स्टार्च का निर्माण नहीं होता है।
– रन्ध्र खुलते समय स्टार्च का शर्करा में परिवर्तन नहीं देखा गया है अपितु स्टार्च से कार्बनिक अम्लों में परिवर्तन देखा गया है।
ग्लाइकोलेट उपापचय परिकल्पना (Glycolate metabolism concept)–
– जेलीट्ज (Zelitch 1963) के अनुसार रक्षक कोशिकाओं में ग्लाइकोलेट का निर्माण रन्ध्र के खुलने में महत्त्वपूर्ण कारक है तथा इसका निर्माण CO2 की न्यून सान्द्रता में होता है। ग्लाइकोलेट से कार्बोहाइड्रेट का निर्माण होता है जिसके फलस्वरूप रक्षक कोशिकाओं का परासरणी दाब बढ़ जाता है। इसके अतिरिक्त इस क्रिया में ATP का निर्माण होता है, जो रन्ध्रों के खुलने व बन्द होने के लिए आवश्यक ऊर्जा का स्रोत है।
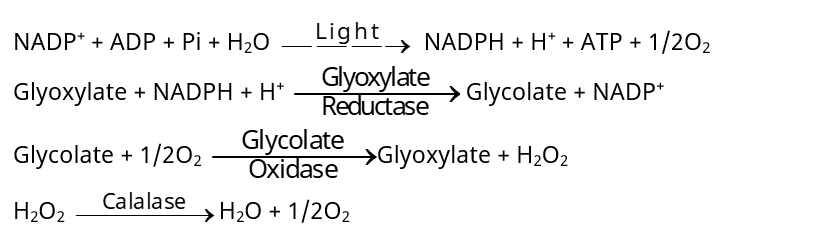
– इस प्रकार ग्लाइकोलेट-ग्लाइआक्सिलेट शटल (Glycolate-glyoxylate shuttle) द्वारा NADPH + H+ का पुन:ऑक्सीकरण होता है तथा ATP का निर्माण होता है। ATP रक्षक कोशिकाओं में जल पम्प करने में सहायक होती है। यह मत अधिकतर वैज्ञानिकों को मान्य नहीं है।
सक्रिय पोटैशियम आयन स्थानान्तरण सिद्धान्त (Active K+ transport theory) –
– यह मत फ्यूजीनो (Fujino, 1967) ने प्रस्तुत किया। फ्यूजीनो के अनुसार रन्ध्रों का खुलना रक्षक कोशिकाओं में K+ की सान्द्रता बढ़ने के कारण होता है। रक्षक कोशिकाओं में K+ आयन एपीडर्मल तथा सहायक कोशिकाओं (subsidiary cells) से प्रवेश करते हैं, जो इनके संग्रहण केन्द्र (store house) का कार्य करती हैं।
– K+ का रक्षक कोशिकाओं में सक्रिय परिवहन ATP के उपयोग से होता है, जो चक्रीय प्रकाश फॉस्फोरिलीकरण (cyclic photophosphorylation) से प्राप्त होती है। इसके अतिरिक्त कुछ ATP श्वसन से भी प्राप्त होती है, जो अन्धकार में रन्ध्रों के खुलने में प्रयुक्त होती है । रन्ध्रों के बन्द होने की प्रक्रिया एब्सिसिक अम्ल (ABA) तथा सम्बन्धित प्रतिरोधकों के नियन्त्रण में होती है।
– प्रकाश की उपस्थिति में रक्षक कोशिकाओं के साइटोप्लाज्म में pH का मान बढ़ जाता है। pH बढ़ने से स्टार्च का अपघटन कार्बनिक अम्ल में होता है। ऐसा माना जाता है कि ग्लाइकोलिसिस द्वारा फॉस्फोइनोलपाइरुवेट (PEP) का निर्माण होता है तथा PEP से आक्जेलो-एसीटिक अम्ल (OAA) का निर्माण होता है, जो शीघ्र ही मैलिक अम्ल (mallic acid) में परिवर्तित हो जाता है। मैलिक अम्ल H+ तथा मैलेट – आयनों में अपघटित हो जाता है। रक्षक कोशिकाओं की प्लाज्माकला पर सक्रिय H+ K+ आयन स्थानान्तरण पम्प (H+ K+ ionexchange pump) की सक्रियता से H+ रक्षक कोशिकाओं से बाहर सहायक कोशिकाओं तथा K+ सहायक कोशिकाओं से रक्षक कोशिकाओं में स्थानान्तरित हो जाते हैं । मैलेट अनायन K+ का आवेश सन्तुलित करने का कार्य करते हैं। रक्षक कोशिकाओं की रिक्तिका में K+ व मैलेट – की सान्द्रता बढ़ने से इनका परासरणी दाब (OP) बढ़ जाता है। रक्षक कोशिकाएँ पास की कोशिकाओं से जल अवशोषित कर स्फीति हो जाती हैं, जिसके फलस्वरूप रन्ध्र खुल जाते हैं।
– अंधकार में रक्षक कोशिकाओं की pH का मान कम हो जाता है, जिससे मैलेट –, H+ से क्रिया कर मौलिक अम्ल का निर्माण करते हैं। इसके साथ ही प्रकाश की अनुपस्थिति में प्रकाश फॉस्फोरिलीकरण बन्द हो जाता हैं तथा ATP संश्लेषण के अभाव में आयन विनिमय पम्प (H+ K+ ion exchange pump) बन्द हो जाता है, जिसके फलस्वरूप K+ का परिवहन सहायक कोशिकाओं में तथा H+ का परिवहन रक्षक कोशिकाओं में होने लगता है। इसके फलस्वरूप रक्षक कोशिकाओं का परासरणी दाब कम हो जाता है तथा जल बहि:परासरण द्वारा पास की कोशिकाओं में स्थानान्तरित हो जाता है। रक्षक कोशिकाएँ शिथिल पड़ जाती हैं, जिसके फलस्वरूप रन्ध्र बंद हो जाते हैं।
– दिन के समय रन्ध्रों का बन्द होना (ABA) एब्सिसिक अम्ल के कारण होता है । CO2 की सान्द्रता पर ABA का निर्माण प्रारम्भ हो जाता है, जो H+ K+ ATP ase विनिमय पम्प को बन्द कर देता है। इसके फलस्वरूप रक्षक कोशिकाओं में उपलब्ध H+ मैलेट से क्रिया कर मैलिक अम्ल का निर्माण कर देते हैं इस प्रकार मैलिक अम्ल से स्टार्च का निर्माण होता है तथा रक्षक कोशिकाओं का परासरणी दाब कम हो जाता है व रन्ध्र बन्द हो जाते हैं।
तमोसक्रिय रन्ध्र गति
(Scotoactive opening & closing of stomata)
– मांसल पौधों (succulents) में रन्ध्र रात्रि में खुले तथा दिन में बन्द रहते हैं।
– इन पादपों में रात्रि के समय कार्बोहाइड्रेट का पूर्ण ऑक्सीकरण नहीं होता, जिसके फलस्वरूप CO2 का निर्माण नहीं हो पाता अपितु कार्बनिक अम्ल बनते हैं।
– चूँकि CO2 का निर्माण नहीं होता इसलिए रात्रि में रन्ध्र खुले रहते हैं। दिन के समय कार्बनिक अम्लों का विघटन होता है जिससे अत्यधिक मात्रा में CO2 का विमोचन होता है, जो रन्ध्रों को बन्द करने के लिए पर्याप्त होती है।
रात्रि के समय (अन्धकार)
2C6H12O6 + 3O2 → 3C4H6O5 + 3H2O
कार्बोहाइड्रेट (मैलिक अम्ल)
दिन के समय (प्रकाश)
C4H6O5 + 3O2 → 4CO2 + 3H2O
– रन्ध्रों के खुलने से पूर्व रक्षक कोशिकाओं में pH का मान O2 की न्यून सान्द्रता के कारण बढ़ जाता है। ऑक्सी श्वसन के अभाव में फासफोइनोल पायरूवेट (PEP) का निर्माण होता है। फास्फोइनोल पायरूवेट कार्बोक्सिलेज (PEP carboxylase.) की सक्रियता द्वारा ओक्जेलेट (OAA) तथा डिहाइड्रोजिनेज द्वारा मैलिक अम्ल में रूपान्तरित हो जाता है।
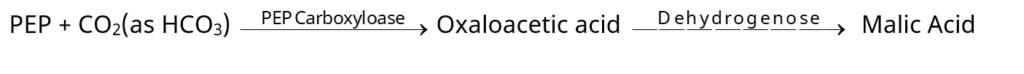
– मैलिक अम्ल का H+ तथा मैलेट – आयनों में अपघटन होता है। H+ सहायक कोशिकाओं तथा K+ रक्षक कोशिकाओं में स्थानान्तरित हो जाते हैं। K+ तथा मैलेट – रक्षक कोशिकाओं की रिक्तिकाओं में स्थानान्तरित होकर परासरणी सान्द्रता बढ़ाते हैं। इसके परिणामस्वरूप रक्षक कोशिकाएँ जल अवशोषित कर स्फीति हो जाती हैं तथा रन्ध्र खुल जाते हैं।
– दिन के समय कार्बनिक अम्लों का विकार्बोक्सिलीकरण (decarboxylation) होता है, जिससे भारी मात्रा में CO2 का निर्माण होता है, फलस्वरूप pH का मान कम हो जाता है इस प्रकार K+ का परिवहन रुक जाता है तथा रक्षक कोशिकाओं का परासरणी दाब कम हो जाने के कारण ये शिथिल हो जाती हैं व रन्ध्र बन्द हो जाते हैं। कार्बनिक अम्लों के विकार्बोक्सिलीकरण से उत्पन्न CO2 रन्ध्रों के बन्द होने पर भी प्रकाश संश्लेषण क्रिया को जारी रखती है।
रन्ध्रों की गति को प्रभावित करने वाले कारक (Factors Affecting Stomatal Movement)
(i) प्रकाश (Light) –
– मांसलोद्भिद मरुद्भिदों के अलावा शेष सभी पादपों में रन्ध्र प्रकाश की उपस्थिति में खुलते हैं तथा अन्धकार में बंद हो जाते हैं। रन्ध्रों के खुलने में नीला व लाल रंग प्रभावी होते हैं।
(ii) ताप (Temperature) –
– लोफ्टफील्ड ने रन्ध्रों के खुलने का ताप गुणांक (Q10) = 2 बताया।
(iii) CO2 सान्द्रता (CO2 – Concentration) –
– पत्तियों की कोशिकाओं में जल की कमी होने पर रन्ध्र CO2 के प्रति संवेदी हो जाते हैं।
– कम CO2 सान्द्रता पर रन्ध्र खुलते हैं जबकि CO2 की अधिक सान्द्रता पर रन्ध्र बंद हो जाते हैं।
(iv) वृद्धि हॉर्मोन (Growth Hormone) –
– साइटोकाइनिन हॉर्मोन रन्ध्र खोलने को प्रेरित करता है।
– K+ आयनों के प्रवेश को बढ़ाकर रन्ध्रों को खुलने के लिए प्रेरित करते हैं।
– हॉर्मोन ABA रन्ध्रों को बंद होने के लिए प्रेरित करते हैं। यह हॉर्मोन साइटोकाइनिन के प्रेरण प्रभाव का विरोध करता है।
– ABA से रक्षक कोशिकाओं की पारगम्यता प्रभावित होती है। यह H+ आयनों के बहिर्वाह (Outflux) को रोक देता है तथा K+ आयनों के बहिर्वाह (Outflux) को बढ़ाता है। इससे रक्षक कोशिकाओं की pH कम हो जाती है। उपर्युक्त प्रभाव CO2 की अधिकता के समय भी पाया जाता है। Cl– आयन भी रंध्रीय गति में सम्मिलित माने जाते हैं।
– उच्च जल तनाव से पत्तियों के हरित लवकों में ABA (Abscisic Acid) बनता है।
(v) वायुमण्डलीय आर्द्रता (Atmospheric Humidity) –
– आर्द्र वातावरण में रन्ध्र अधिक तक व अधिक मात्रा में खुले रहते हैं जबकि शुष्क वातावरण में रन्ध्र बंद हो जाते हैं या कम खुले रहते हैं। उच्च वायु आर्द्रता पर वाष्पोत्सर्जन पूर्णत: बंद, किन्तु रन्ध्र पूर्ण: खुले रहते हैं।
वाष्पोत्सर्जन को प्रभावित करने वाले कारक
(Factors Affecting the Rate of Transpiration)
– ये दो प्रकार के होते हैं-
(i) बाह्य कारक या वातावरणीय कारक
(ii) वायुमण्डलीय आर्द्रता (Atmospheric Humidity)

– कम वायुमण्डलीय आर्द्रता पर वायुमण्डल शुष्क होता है इसलिए वाष्पोत्सर्जन अधिक तेज होता है, जबकि उच्च वायुमण्डलीय आर्द्रता पर वायुमण्डल नम रहता है, जिसके फलस्वरूप वाष्पोत्सर्जन की दर घट जाती है। रन्ध्र पूर्णत: खुले रहते हैं।
– इसी कारण गर्मियों में वाष्पोत्सर्जन की दर बढ़ जाती है तथा बरसात के दिनों में वाष्पोत्सर्जन घट जाता है।
ताप (Temperature) – Tr ∝ Temperature
– वाष्पोत्सर्जन के Q10 का मान 2 होता है अर्थात् ताप में 10oC की वृद्धि करने पर वाष्पोत्सर्जन की दर लगभग दुगुनी हो जाती है। (Loftfield ने बताया)
– उच्च ताप पर वायु की जल वाष्प ग्रहण क्षमता बढ़ जाती है जिसके फलस्वरूप वाष्पोत्सर्जन की दर बढ़ जाती है। (क्योंकि Vapour Pressure कम होता है।)
– इसके विपरीत कम ताप पर वायु की जल वाष्प ग्रहण करने की क्षमता घट जाती है, जिसके कारण वाष्पोत्सर्जन घट जाता है।
प्रकाश (Light) –
– वाष्पोत्सर्जन प्रकाश द्वारा प्रेरित होता है। यह ताप वृद्धि भी करता है।
– वाष्पोत्सर्जन का क्रिया स्पैक्ट्रम (Action-spectrum) नीला तथा लाल होता है।
– नीले प्रकाश में वाष्पोत्सर्जन की क्रिया लाल प्रकाश की तुलना में अधिक तीव्र होती है, क्योंकि नीले प्रकाश में रन्ध्र अपनी पूरी क्षमता में खुले रहते हैं।
वायुमण्डलीय दाब (Atmospheric Pressure) –
– कम वायुमण्डलीय दाब पर वायु विरल हो जाती है व इसकी गति बढ़ जाती है। इससे विसरण की दर बढ़कर वाष्पोत्सर्जन की दर को बढ़ा देती है। किसी पौधे को धरातल से ऊपरी स्थानों पर ले जाने पर वाष्पोत्सर्जन बढ़ जाता है।
आन्तरिक कारक (Internal Factors) –
– ये कारक पादप संरचना से संबंधित होते हैं।
वाष्पोत्सर्जन क्षेत्रफल (Transpiring Area) –
– छँटाई-कटाई (Prunning) से वाष्पोत्सर्जन तीव्रता बढ़ेगी किन्तु कुल वाष्पोत्सर्जन तो कम हो जाएगा।
– पर्ण की संरचनात्मक विशेषताएँ – उपत्वचा (Cuticle) – उपत्वचा के मोटी होने से वाष्पोत्सर्जन में कमी होती है।
रन्ध्र विशेषताएँ –
– रन्ध्रों की संरचना, रन्ध्रों की स्थिति, रन्ध्रों के बीच की दूरी, रन्ध्रों की इकाई क्षेत्र में संख्या तथा रन्ध्रों की सक्रियता से वाष्पोत्सर्जन प्रभावित होता है।
– रन्ध्रीय सूचकांक (Stomatal Index) 𝐼=𝑆𝐸+𝑆
(Salisbury द्वारा)
I = Stomatal Index
S = रन्ध्रों की संख्या/इकाई क्षेत्रफल
E = एपीडर्मल कोशिकाओं की संख्या इकाई क्षेत्रफल में
– धँसे हुए रन्ध्रों से (कैर), धँसी हुई बाह्य त्वचा पर रोम होने से (कनेर) पास-पास रन्ध्र होने से इकाई क्षेत्र में रन्ध्रों की संख्या कम होने से, रन्ध्र कम मात्रा में वे कम समय तक खुलने से वाष्पोत्सर्जन में कमी होती है।
वाष्पोत्सर्जन का महत्त्व (Significance of Transpiration)
– ताप का नियमन (Regulation of Temp.) – वाष्पोत्सर्जन की क्रिया के कारण पत्तियों की सतह पर शीतलन प्रभाव (Cooling Effect) उत्पन्न होता है। जिसके कारण पौधों का निश्चित ताप बना रहता है।
– पादप शरीर में अवशोषित खनिजों का स्थानांतरण
– निष्क्रिय जल अवशोषण (Water Absorption)
– रसारोहण (Ascent of Sap)
– खनिजों का वितरण (Distribution of Salts absorbed)
– Gaseous Exchange में।
– प्रकृति में जल चक्र का नियंत्रण
बिंदु स्राव (Guttation) –

– पादप के अक्षतिग्रस्त (Uninjured) भाग या पत्तियों के शिरा अंतिमों (Veinlets) तथा किनारों से जल की बूँदों के रूप में हानि बिन्दु स्राव कहलाती है।
– बिंदु स्राव की क्रिया शाकीय पादपों में पाई जाती है। जल अवशोषण अधिक व वाष्पोत्सर्जन न्यून होने पर।
– बिंदु स्राव में निकलने वाले द्रव्य में जल के अतिरिक्त कुछ कार्बनिक तथा अकार्बनिक पदार्थ घुले हुए रहते हैं अर्थात् यह शुद्ध जल नहीं होता।
– बिंदु स्राव पत्तियों के किनारों पर शिरिकाओं के स्थान पर पाए जाने वाले विशेष छिद्रों (जो सदैव खुले) द्वारा होता है, जिन्हें जलरन्ध्र (Hydathodes or Water Stomata) कहते हैं। बिंदु स्राव प्राय: रात्रि व प्रात: काल होता है।
– प्रत्येक जलरन्ध्र रंगहीन अधिचर्मीय कोशिकाओं द्वारा बनता है।
– जलरन्ध्र के नीचे मृदुतकीय एवं ढीला (Loose) ऊतक स्थित होता है जिसे ऐपीथेम (Epithem) कहते हैं। इसे ट्रांसफर ऊतक भी कहते हैं।
– बिंदु स्राव की क्रिया मूलदाब (Root Pressure) के कारण होती है। जो वल्कुट कोशिकाओं में उत्पन्न होता है।
रस स्त्रावण (Bleeding) –
– पादप के कटे हुए या क्षतिग्रस्त भाग से तरल का स्रावण होना रस स्रावण (Exudation) या Bleeding कहलाता है।
– यह क्रिया उच्च मूलदाब के कारण होती है। रबर व अफीम इसी प्रक्रिया से प्राप्त किए जाते हैं।
– इस क्रिया द्वारा ‘शुगर मैपल’ से शर्करा प्राप्त की जाती है।
म्लानि (Wilting) –
– म्लानि (Wilting) पौधों के मुलायम भागों में कोशिका की Turgidity की कमी से मुरझाने के लक्षण उत्पन्न हो जाते हैं, इसी घटना को म्लानि कहा जाता है जो प्राय: दोपहर में अधिक वाष्पोत्सर्जन या मृदा में जल की कमी से देखी जाती है। दीर्घकालीन सूखे के समय भी पौधों में ऐसा होता है।
– जब पौधों को पर्याप्त जल उपलब्ध होने पर म्लानि के लक्षण समाप्त नहीं होते तो इसे ‘स्थाई म्लानि’ कहा जाता है।
फ्लोएम द्वारा पादपों में परिवहन–
– फ्लोएम द्वारा कार्बनिक विलेयों के अभिगमन अथवा स्थानान्तरण को स्पष्ट करने के लिए अनेक मत प्रस्तुत किए गए हैं। अभी तक प्रस्तुत कोई भी क्रियाविधि पदार्थों के स्थानान्तरण को संतोषजनक ढंग से स्पष्ट करने में असमर्थ है। इस सम्बन्ध में प्रस्तुत कुछ मत इस प्रकार हैं–
1. विसरण परिकल्पना (Diffusion hypothesis) –
– इस मत के अनुसार कार्बनिक पदार्थों का स्थानान्तरण इनकी अधिक सान्द्रता से कम सान्द्रता क्षेत्र की ओर होता है। इस प्रकार इनका स्थानान्तरण संग्रहण क्षेत्र (पत्तियों) से उपभोग क्षेत्र (जड़ व अन्य भागों) की ओर होता है क्योंकि पत्तियों में संश्लेषण के कारण इनकी सान्द्रता अधिक होती है तथा जड़ व अन्य भागों में उपभोग के कारण इनकी न्यून सान्द्रता पाई जाती है।
– विसरण परिकल्पना को पूर्ण रूप से नकार दिया गया है क्योंकि कार्बनिक पदार्थों के परिवहन की वास्तविक दर विसरण दर से हजारों गुणा अधिक होती है। फ्लोएम में परिवहन उपापचयी क्रियाओं पर निर्भर करता है जबकि विसरण एक भौतिक क्रिया है।
2. जीवद्रव्य प्रवाह गति परिकल्पना (Protoplasmic streaming hypothesis) –

– इस परिकल्पना को डी-व्रीज (De Vries, 1885) ने प्रस्तावित किया तथा कर्टिस (Curtis, 1935) ने विकसित किया था। इस परिकल्पना के अनुसार विलेय कणों का परिवहन फ्लोएम में विसरण (diffusion) तथा जीवद्रव्य प्रवाह (streaming) क्रियाओं का परिणाम है। कार्बनिक पदार्थ चालनी नलिकाओं में एक सिरे से दूसरे सिरे तक जीवद्रव्य प्रवाही गति द्वारा गमन करते हैं तथा एक चालनी नलिका से दूसरी चालनी नलिका में रन्ध्रों से विसरण द्वारा परिवहन होता है तथा कोशिका द्रव्य की प्रवाह गति विसरण को त्वरित करती है। जीवद्रव्य प्रवाही गति के कारण एक ही चालनी नलिका में अलग-अलग प्रवणता वाले दो पदार्थ एक-दूसरे के विपरीत दिशाओं में साथ-साथ परिवहन कर सकते हैं।
प्रमाण (Evidences) –
– चालनी नलिकाओं में जीवद्रव्य प्रवाह गति प्रमाणित हो चुकी है।
– यह परिकल्पना एक ही चालनी नलिका में दोतरफा परिवहन (bidirectional movement) को स्पष्ट करती है।
– यह परिकल्पना कणों के स्थानान्तरण तथा कोशिका द्रव्य प्रवाह पर ताप, ऑक्सीजन तथा श्वसन निरोधी यौगिकों के प्रभाव को भी स्पष्ट करती है।
आपेक्ष (Objection) –
– जीवद्रव्य प्रवाह गति केवल युवा चालनी नलिकाओं में पाई जाती है, परिपक्व चालनी नलिकाओं में जीवद्रव्य प्रवाह गति का अभाव होता है जबकि वृक्षों में युवा चालनी नलिकाओं की अपेक्षा परिपक्व चालनी नलिकाओं की संख्या अधिक होती है।
– जीवद्रव्य प्रवाह गति की दर 2-5 सेंटीमीटर प्रति घण्टा होती है, जबकि कार्बनिक अणुओं का परिवहन 100 से 150 सेंटीमीटर प्रति घन्टा की दर से होता है।
– परिपक्व चालनी नलिकाओं में जीवद्रव्य अल्प मात्रा में होता है।
3. पारकोशिकीय प्रवाही परिकल्पना (Transcelluar streaming)–
– थैने (Thaine, 1964) ने कोशिका द्रव्य प्रवाही परिकल्पना को रूपान्तरित किया। थैने के अनुसार चालनी नलिकाओं में प्रोटीनी नलिकाकार तन्तुगुच्छ अथवा स्ट्रैण्ड्स (tubular strands) पाए जाते हैं जो चालनी रन्ध्रों से होकर एक चालनी कोशिका से दूसरी चालनी कोशिका तक निरन्तर होते हैं। कार्बनिक पदार्थों का परिवहन इन स्ट्रेण्ड्स के माध्यम से क्रमाकुंचन (peristaltic movement) द्वारा होता है। एक चालनी नलिका में अनेक स्ट्रेण्ड की उपस्थिति के कारण दोनों दिशाओं में कार्बनिक पदार्थों के परिचालन की व्याख्या भी की जा सकती है। इस परिकल्पना को मान्यता प्राप्त नहीं है क्योंकि इन स्ट्रेण्ड्स की उपस्थिति इलेक्ट्रॉन सूक्ष्मदर्शी द्वारा प्रमाणित नहीं हुई है।
4. वैद्युत-परासरण परिकल्पना

– यह परिकल्पना फेन्सम (Fensom,1957) तथा स्पैनर (Spanmer,1958) द्वारा स्वतन्त्र रूप से प्रस्तावित की गई थी। इस मत के अनुसार चालनी नलिका में चालनी पट्टिका पर विद्युत विभव उपस्थित होता है जो विलेय कणों के परिवहन के लिए उत्तरदायी होता है।
– चालनी नलिकाओं में जल तथा विलेय कणों का संचलन विद्युत प्रवणता के अनुरूप अधिक धनावेश से कम धनावेश के क्षेत्र की दिशा में होता है। विद्युत प्रवणता को बनाए रखने का कार्य K+ के परिसंचरण (circulation) द्वारा होता है। K+ आयनों की सान्द्रता चालनी पट्टिका के ऊपर की ओर अधिक तथा नीचे की ओर कम होती है, इस प्रकार विलेय अणु K+ के साथ विद्युत प्रवणता की दिशा में ऊपर से नीचे की ओर गति करते हैं। विलेय तथा K+ आयनों के पट्टिका के नीचे पहुँचने पर पट्टिका के नीचे K+ आयन सान्द्रता बढ़ने लगती है तो सहकोशिकाएँ K+ आयनों को अवशोषित कर पट्टिका को ऊपर की ओर पम्प कर देती हैं। इस प्रकार K+ आयनों का यह चक्रीय प्रवाह बना रहता है। पोटैशियम आयनों के इस चक्रीय प्रवाह के लिए आवश्यक ऊर्जा (ATP) सहचर कोशिकाओं से प्राप्त होती है।
– प्रमाण (Evidences) –
– पौधे में पोटैशियम की न्यूनता होने पर स्थानान्तरण दर में भी कमी आ जाती है।
– चालनी पट्टिका के दोनों ओर विभवान्तर की पुष्टि की गई है।
5. द्रव्यमान प्रवाह परिकल्पना अथवा मुन्च परिकल्पना (Mass flow hypothesis or Munch hypothesis) –
– यह परिकल्पना मुन्च (Munch, 1930) द्वारा प्रस्तुत की गई थी, इसे मुन्च परिकल्पना भी कहते हैं। इस परिकल्पना के अनुसार कार्बनिक पदार्थों का परिवहन द्रव्य (mass) के रूप में होता है। उत्पादन व उपभोग अंगों के मध्य स्फीति दाब-प्रवणता (turgor pressure-gradient) इस परिवहन के लिए उत्तरदायी होती है।
– मुन्च के अनुसार फ्लोएम की चालनी नलिकाएँ सिरों से आपस में जुड़ी रहती हैं तथा इनका जीवद्रव्य प्लाज्मोडेस्माटा (plasmodesmata) द्वारा जुड़ा रहता है जिससे एक सतत तन्त्र का निर्माण होता है। इसे सिम्प्लास्ट (symplast) कहते हैं, जो पादप के शीर्ष से आधार तक सतत होता है तथा सीधे अक्ष की दिशा में पारगम्य (permeable) होता है।
– पत्तियों में (स्रोत) प्रकाश-संश्लेषण के फलस्वरूप कार्बोहाइड्रेट्स का निर्माण होता है जो पर्णमध्योत्तक कोशिकाओं में सूक्रोज (शर्करा) के रूप में एकत्र होता है। पर्णमध्योतक कोशिकाओं में सूक्रोज के एकत्र होने पर इनका परासरणी दाब बढ़ जाता है तथा ये जाइलम तत्त्वों से जल ग्रहण करती है जिससे इनका स्फीति दाब (turgor pressure) बढ़ जाता है। उच्च स्फीति दाब के कारण विलयन इन कोशिकाओं से चालनी नलिकाओं में प्रवेश कर जाता है। पर्णमध्योत्तक कोशिकाओं में प्रकाश संश्लेषण के फलस्वरूप शर्करा की सान्द्रता लगातार बनी रहती है जिसके कारण इनका स्फित दाब भी बना रहता है। इसके विपरीत जड़ या उपयोग केन्द्र (सिंक) में शर्करा या कार्बनिक पदार्थों का उपभोग अथवा अघुलनशील रूप में (स्टार्च) संग्रहण होता रहता है जिससे इनका परासरणी दाब कम हो जाता है अर्थात् स्फीति दाब (turgor pressure) कम रहता है। इस प्रकार स्रोत (पर्णमध्योतक) तथा उपभोग सिरों (जड़ या अन्य भाग) के मध्य स्फीति दाब प्रवणता (turgor pressure gradient) उत्पन्न हो जाती है। फ्लोएम के दोनों सिरों के मध्य उत्पन्न स्फीति दाब प्रवणता के फलस्वरूप विलयन (घुलनशीन विलेय + जल) का द्रव्य प्रवाह (mass flow) पत्तियों से जड़ की ओर फ्लोएम में होता रहता है। जड़ों या उपभोग वाले सिरे पर शर्करा का उपभोग हो जाता है या फिर स्टार्च के रूप में संचित हो जाता है तथा जल जाइलम वाहिकाओं में विसरित हो जाता है तथा ऊपर को परिवहन करता है।
एन्जाइम
– कोशिकाओं में विभिन्न जैव रासायनिक क्रियाएँ कुछ अति सक्रिय कार्बनिक पदार्थों की अल्प मात्रा की उपस्थिति में सम्पन्न होती है। ये पदार्थ जैव उत्प्रेरकों (Biocatalysts) की भाँति व्यवहार करते हैं, इन्हें एन्जाइम अथवा प्रकिण्व कहते हैं। एन्जाइम रासायनिक अभिक्रियाओं की दर को अभिकारकों (Reactants) की सक्रियण ऊर्जा कम करके बढ़ा देते हैं।
– कोशिकाओं में कुछ अभिक्रियाएँ संश्लेषण प्रकार की होती हैं। जिनमें ऊर्जा की आवश्यकता होती है इन्हें उपचय अभिक्रियाएँ (Anabolic processes) कहते हैं जबकि कुछ अभिक्रियाएँ विघटन प्रकार की होती हैं जिनमें ऊर्जा मुक्त होती हैं, इन्हें अपचय अभिक्रिया (Catabolic processes) कहा जाता है।
– कोशिकाओं में ये दोनों अभिक्रियाएँ मिलकर उपापचय पथों का निर्माण करती हैं। एन्जाइम की सर्वस्वीकृत परिभाषा है- ‘प्रोटीन प्रकृति के ऐसे पदार्थ जो जीवित कोशिकाओं में जैव उत्प्रेरक का कार्य करते हैं, एन्जाइम कहलाते हैं।’
– एन्जाइम की जैव उत्प्रेरक के रूप में पहचान बर्जिलियस (Bergelius, 1835) ने की। एडवर्ड बुकनर (Edward Buchner) ने सर्वप्रथम यीस्ट की कोशिकाओं से जायमेज (Zymase) नामक एन्जाइम जटिल को खोजा।
– एन्जाइम शब्द का सर्वप्रथम प्रयोग विली कुहने (Willy Kuhne, 1878) ने यीस्ट विलयन के किण्वन के लिए किया।
– जे.बी. सुमनर (J.B. Sumner, 1926) ने सर्वप्रथम यूरिऐज एन्जाइम का विशुद्ध क्रिस्टलीय रूप प्राप्त किया। सुमनर एवं नॉरथ्रोप (Sumner and Northrope, 1930) ने सप्रमाण स्थापित किया कि समस्त एन्जाइम्स रासायनिक दृष्टि से प्रोटीन होते हैं।
– अप्रोटीनी एन्जाइम (RNA के उत्प्रेरकीय अणु) की खोज सर्वप्रथम सेक एवं अल्टमेन (Thomas Cech and Altman) द्वारा की गई जिसे राइबोजाइम (Ribozyme) कहा गया है।
एन्जाइम की संरचना –
– प्रोस्थेटिक समूह (Prosthetic group) – जब कार्बनिक प्रकृति का अप्रोटीन भाग एपोएन्जाइम से सघनता से एवं दृढ़तापूर्वक अनुबंधित रहता है, उसे प्रोस्थेटिक समूह कहते हैं। उदाहरण – साइटोक्रोम, फ्लेवो प्रोटीन
– सह एन्जाइम (Co-enzyme) – जब अप्रोटीन भाग एपोएन्जाइम से ढीले अथवा आसानी से पृथक् होने योग्य तथा पुनः संलग्न होने योग्य रूप में अनुबंधित रहता है तो इस भाग को सहएन्जाइम या को-एन्जाइम कहते हैं। जैसे- NAD, NADP, FAD, Co-A आदि।
– सक्रियक (Activator) – जब अप्रोटीन भाग अकार्बनिक प्रकृति का कोई धातु आयन हो तो उसे सक्रियक कहते हैं। जैसे- Fe
होलो एन्जाइम = एपो एन्जाइम + अप्रोटीन भाग (कोफेक्टर)
(i) प्रोस्थेटिक समूह
(ii) सह-एन्जाइम
(iii) सक्रियक
– होलो एन्जाइम से सह-एन्जाइम को डायलेसिस की प्रक्रिया द्वारा आसानी से पृथक् किया जा सकता है।
एन्जाइमों के विशिष्ट गुण (Specific Properties of Enzymes)
– प्रोटीन प्रकृति (Protein nature) – सभी एन्जाइम रासायनिक रूप से प्रोटीन होते हैं। इनमें प्रोटीन के अतिरिक्त कभी-कभी अकार्बनिक अथवा कार्बनिक पदार्थ के परमाणु, आयन अथवा समूह भी उपस्थित हो सकते हैं।
– कोलॉइडी प्रकृति (Colloidal nature) – समस्त एन्जाइम कोलॉइडी प्रकृति के होते हैं। इनका पृष्ठीय क्षेत्रफल अधिक होने के कारण ये जैव रासायनिक अभिक्रियाओं के लिए वृहत् सतह प्रदान करते हैं, जिससे अभिक्रियाएँ तीव्रदर से सम्पन्न होती हैं।
– विशिष्टता (Specificity) – सभी एन्जाइम अपनी अभिक्रिया के प्रति विशेषता रखते हैं। सामान्यतः एक एन्जाइम केवल एक ही अभिक्रिया को उत्प्रेरित करता है या कुछ विशेष प्रकार के रासायनिक आबंधों पर क्रियाशील रहता है जबकि कुछ ही एन्जाइम ऐसे हैं जो दो या दो से अधिक भिन्न प्रकार की अभिक्रियाओं को भी उत्प्रेरित करते हैं।
– एन्जाइम विशिष्टता सामान्यत: चार प्रकार की होती है –
1. निरपेक्ष विशिष्टता (Absolute specificity) – इस प्रकार की विशिष्टता वाले एन्जाइम केवल एक प्रकार की अभिक्रिया को ही उत्प्रेरित करते हैं। जैसे यूरिएज द्वारा केवल विशिष्ट यूरिया का जल अपघटन किया जाता है।
2. समूह विशिष्टता (Group specificity) – कुछ एन्जाइम सम्बन्धित क्रियाकारक (Substrate) के समूह विशेष पर ही उत्प्रेरण करते हैं। जैसे हेक्सोकाइनेज विभिन्न प्रकार की हेक्सोज शर्कराओं पर क्रियाशील होते हैं।
3. बन्ध विशिष्टता (Bond specificity) – कुछ एन्जाइम एक विशेष प्रकार के रासायनिक बंध की ही पहचान कर इस पर क्रियाशीलता दर्शाते हैं जैसे राइबोन्यूक्लिएज एन्जाइम द्वारा पिरिमिडिन क्षारक के 3′ (3 prime) सिरे पर ही फोस्फोडाइएस्टर बन्ध का जल अपघटन किया जाता है।
4. त्रिविम विशिष्टता (Stereo specificity) – कुछ एन्जाइम क्रियाधारों के केवल विशेष प्रकार के त्रिविम समावयवी पर ही क्रियाशील होते हैं, दूसरे रूपों पर नहीं। जैसे- अमीनो अम्ल ऑक्सीडेज के L- (levo) अमीनो अम्ल द्वारा होने वाली अभिक्रियाओं को ही उत्प्रेरित करते हैं, D अमीनो अम्ल पर नहीं।
एन्जाइम क्रिया को प्रभावित करने वाले कारक (Factors affecting Enzymatic Action)
– तापक्रम (Temperature) – एन्जाइम क्रिया के लिए अनुकूल ताप 20°C से 35°C तक होता है। इस सीमा में 10°C तापक्रम में वृद्धि करने पर अभिक्रिया की दर 2 से 2½ गुना तक बढ़ जाती है। 35°C से अधिक तापक्रम पर तापीय विकृतीकरण के कारण अभिक्रिया की गति मंद हो जाती है व अति उच्च तापक्रम पर बंद हो जाती है।
– pH मान – अधिकांश एन्जाइम 5.0 से 7.5 pH की परास में दक्षतापूर्वक कार्य करते हैं। pH के मान में कमी अथवा अधिकता एन्जाइम की क्रियाशीलता को कम करती है।
– क्रियाधारों की सान्द्रता (Concentration of substrate) – निम्न सान्द्रता पर एन्जाइम क्रिया की गति कम हो जाती है क्योंकि एन्जाइम अणु की सतह पर स्थित सभी सक्रिय स्थलों पर क्रियाधार अणुओं का संयोग नहीं होता है। क्रियाधार की सान्द्रता बढ़ाने पर क्रिया की गति भी बढ़ती है। एक निश्चित सान्द्रता से अधिक सान्द्रता पर क्रिया की गति स्थिर हो जाती है। पर्याप्त सान्द्रता पर होने वाली अधिकतम गति को अधिकतम वेग कहते हैं।
– एन्जाइम सान्द्रता (Enzyme concentration) – क्रियाधार की उपयुक्त सान्द्रता के रहते एन्जाइम अभिक्रिया की गति एन्जाइम सान्द्रता के अनुक्रमानुपाती होती है। एन्जाइम सान्द्रता के बढ़ाने पर अभिक्रिया की गति धीरे-धीरे बढ़ती है तथा क्रियाधारों की सान्द्रता के सीमाकारी प्रभाव के कारण किसी एक बिन्दु पर रुक कर स्थिर हो जाती है। अतः एन्जाइम सान्द्रता बढ़ाने के साथ-साथ क्रियाधार की सान्द्रता बढ़ाने पर अभिक्रिया की गति बढ़ जाती है।
– अन्तिम उत्पाद की सान्द्रता (Concentration of end product) – अन्तिम उत्पाद की सान्द्रता में वृद्धि के कारण एन्जाइम उत्प्रेरित क्रिया में मन्दता आती है। यह मन्दता बढ़ते-बढ़ते साम्यावस्था पर क्रिया को बिल्कुल शून्य कर देती है तथा साम्यावस्था के पश्चात् क्रिया विपरीत दिशा में प्रारम्भ हो जाती है।
– एन्जाइम निरोधक एवं विष (Enzyme inhibitors and poisons) – वे पदार्थ जो एन्जाइम की उत्प्रेरण क्षमता को कम कर देते हैं, उन्हें एन्जाइम निरोधक कहते हैं। ये निरोधक पदार्थ सामान्यतः एन्जाइम के सक्रिय स्थलों से संयोग करके उन्हें निष्क्रिय कर देते हैं।
– संदमन अभिक्रिया दो प्रकार की होती है–
– प्रतिस्पर्धी निरोधक (Competitive inhibition) – ऐसे निरोधक पदार्थों की संरचना क्रियाधार अणुओं से मिलती-जुलती होती है। अतः ये पदार्थ एन्जाइम अणुओं के सक्रिय स्थलों से जुड़ने में क्रियाधार अणुओं से प्रतिस्पर्धा रखते हैं जिससे एन्जाइम की क्रिया मन्द हो जाती है। जैसे- मेलॉनिक अम्ल सक्सीनिक अम्ल का प्रतिस्पर्धी निरोधक है। इस प्रकार के संदमन को क्रियाधार की सान्द्रता में वृद्धि करके रोका जा सकता है।
– अप्रतिस्पर्धी निरोधक (Non-Competitive inhibition) – इस प्रकार के निरोधक पदार्थ स्थायी रूप से एन्जाइम अणुओं से जुड़कर उनमें संरचनात्मक परिवर्तन ला देते हैं। यह स्थायी संयोजन एन्जाइम अणु को विकृत कर देता है। ऐसे पदार्थों को कोशिका आविष कहा जाता है, जैसे– Pb++, Hg++, Ag++ आदि आयन। सायनाइड श्वसन क्रिया में साइटोक्रोम ऑक्सीडेज को संदमित कर देता है। इस प्रकार के संदमन को एन्जाइम की सान्द्रता बढ़ाकर रोका जा सकता है।
एन्जाइम क्रिया की विधि (Mode of enzyme action)
– समस्त एन्जाइमों की कार्यप्रणाली एन्जाइम व अभिक्रिया में भाग लेने वाले क्रियाधारों की प्रकृति पर निर्भर करती है। एन्जाइम अभिक्रिया का आरम्भन व अभिक्रिया की दर एन्जाइम क्रियाधार जटिल (Enzyme substrate complex) के बनाने व अभिक्रिया के प्रारम्भ होने के लिए आवश्यक सक्रियण ऊर्जा में कमी करने से संबंधित है। अतः एन्जाइम की कार्यप्रणाली को निम्नलिखित प्रकार से समझा जा सकता है–
1. एन्जाइम क्रियाधार जटिल का निर्माण
2. सक्रियण ऊर्जा में कमी
1. एन्जाइम क्रियाधार जटिल या संकुल का निर्माण (Formation of enzyme substrate complex, ESC) – सभी प्रकार की एन्जाइम उत्प्रेरित अभिक्रियाओं में सबसे पहले एन्जाइम क्रियाधारों से मिलकर एक अस्थायी यौगिक का निर्माण करते हैं जिसे एन्जाइम-क्रियाधार जटिल या संकुल (Enzyme substrate complex) कहते हैं। यह संकुल विखण्डित होकर उत्पाद बनाता है व एन्जाइम को पृथक् कर देता है।
क्रियाधार + एन्जाइम à एन्जाइम क्रियाधार जटिल (अस्थायी) एन्जाइम क्रियाधार जटिल à एन्जाइम + उत्पाद
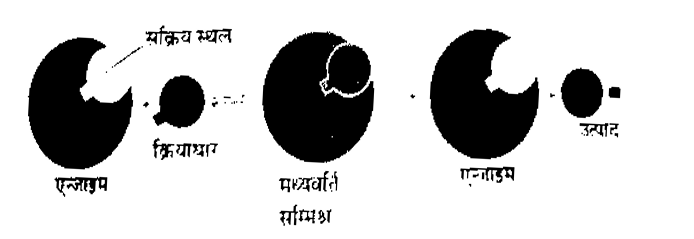
– प्रत्येक एन्जाइम की सतह पर कोई विशेष स्थान (Specific site) होता है जहाँ पर क्रियाकारक जुड़ सकते हैं। उन स्थानों को सक्रिय स्थल (Active sites) कहते हैं। एन्जाइम में सक्रिय स्थल इतने पास-पास होते हैं कि क्रियाधार यहाँ जुड़कर आसानी से क्रिया कर सकते हैं। जब क्रियाधार एन्जाइम सतह से जुड़ते हैं, तब एन्जाइम का आकार परिवर्तित हो जाता है व क्रियाधारों के मध्य नये बन्ध बनते हैं। इन बन्धों के कारण बनने वाला उत्पाद एन्जाइम से पृथक् हो जाता है एन्जाइम मुक्त होकर पुनः क्रियाधारों के अन्य अणुओं से जुड़ जाते हैं एवं नए एन्जाइम क्रियाधार संकुल का निर्माण करते हैं।
– एन्जाइम क्रियाधार संकुल बनने की प्रक्रिया को निम्नलिखित दो सिद्धान्तों से समझाया जा सकता है–
(i) ताला कुंजी सिद्धान्त (Lock and Key Theory) – इस सिद्धान्त का प्रतिपादन एमिल फिशर (Emil Fischer, 1984) द्वारा किया गया था। इस सिद्धान्त के अनुसार एक ताला उसी की चाबी से ही खोला जा सकता है, ठीक इसी प्रकार एक विशेष आकार का क्रियाधार ही एन्जाइम की विशेष सक्रिय सतह से बंध बना सकता है। अत: इस प्रकार की एन्जाइम द्वारा उत्प्रेरित होने वाली अभिक्रिया में विशेष प्रकार के क्रियाधार ही विशेष एन्जाइम के सक्रिय स्थल पर जुड़ सकते हैं।
(ii) प्रेरित आसंजन सिद्धान्त (Induced fit Theory) – इस सिद्धान्त का प्रतिपादन कोशलैण्ड (Koshland, 1966) द्वारा किया गया था। इस सिद्धान्त के अनुसार एन्जाइम में पाए जाने वाले सक्रिय स्थल दृढ़ नहीं होते हैं अर्थात् ये सक्रिय स्थल प्रारम्भ से क्रियाधार अणु की संरचना के पूरक नहीं होते हैं, लेकिन क्रियाधार जैसे ही एन्जाइम के सम्पर्क में आते हैं, सक्रिय स्थल उसके अनुपूरक हो जाते हैं। इस सिद्धान्त के अनुसार एन्जाइम में उपस्थित सक्रिय स्थल क्रियाधारों द्वारा प्रेरित होकर अनुपूरक बनते हैं। इस कारण यह सिद्धान्त प्रेरित आसंजन सिद्धान्त कहलाता है। सरल भाषा में यह कहा जा सकता है कि एन्जाइम का क्रियास्थल ताला चाबी सिद्धान्त द्वारा प्रतिपादित विचार के अनुसार निश्चित या दृढ़ नहीं होकर परिवर्तनशील होता है। अतः क्रियाधार का सम्पर्क होने पर इसे संयोजित करने के लिए अपनी संरचना में मामूली परिवर्तन कर लेते हैं जिससे एन्जाइम सब्सट्रेट कॉम्पलेक्स(एन्जाइम क्रियाधार जटिल) बनता है।
2. सक्रियण ऊर्जा में कमी करना (Lowering of Activation Energy) – रासायनिक अभिक्रियाओं में ऊर्जा मुक्त होती है, लेकिन इन अभिक्रियाओं को आरम्भ न करने के लिए भी कुछ ऊर्जा की आवश्यकता होती है। अभिक्रिया आरम्भ के लिए आवश्यक ऊर्जा, सक्रियण ऊर्जा (Activation energy) कहलाती है। एन्जाइमों में यह विशेषता होती है कि वे क्रियाधार के अणुओं के सक्रियण के लिए आवश्यक ऊर्जा को कम कर देते हैं। अत: एन्जाइम की उपस्थिति में क्रियाकारक कम सक्रियण ऊर्जा पर ही क्रियाफलों में परिवर्तित हो जाते हैं।

एन्जाइमों की नामकरण पद्धति (Nomenclature of Enzymes)
– एन्जाइम नामकरण की दो प्रचलित पद्धतियाँ हैं–
1. क्रियाधार के आधार पर (On the basis of substrate) – इस पद्धति के अनुसार क्रियाधार के नाम के अन्त में ऐज लगा देने से उस एन्जाइम का नाम बन जाता है जो इस क्रियाधार पर आक्रमण करता है जैसे सुक्रेज, माल्टेज, लाइपेज जो क्रमशः शर्करा, माल्ट (कार्बोइड्रेट)तथा लिपिइड्स (वसाएँ) पर क्रियाशील होते हैं।
2. रासायनिक क्रिया के आधार पर (On the basis of chemical reaction) – यह पद्धति अधिक लोकप्रिय एवं व्यवहार में है। इस पद्धति के अंतर्गत एन्जाइमों का नाम उनके द्वारा उत्प्रेरित क्रिया के आधार पर रखा जाता है। उदाहरण के लिए हाइड्रोलेज (Hydrolase), ऑक्सीडेज (Oxydase), कार्बोक्सीलेज (Carboxyloe), डिहाइड्रोजिनेज (Dehydrogenase)। कई एन्जाइमों का नामकरण उनके द्वारा उत्प्रेरित होने वाले क्रियाधार व अभिक्रिया की प्रकृति दोनों के आधार पर किया जाता है। जैसे- L ग्लूटेमिक डिहाइड्रोजिनेज इसमें L– ग्लूटेमिक अम्ल क्रिया कारक है तथा इसके द्वारा उत्प्रेरित होने वाली अभिक्रिया डिहाड्रोजनीकरण है।
एन्जाइम्स का वर्गीकरण (Classification of Enzymes)
– ऑक्सीडोरिडक्टेजेज (Oxidoreductases) – ये एन्जाइम अपने क्रियाधारों में से इलेक्ट्रॉन अथवा हाइड्रोजन हटाकर अथवा जोड़कर उनका ऑक्सीकरण अथवा अपचयन करते हैं।
जैसे – साइटोक्रोम ऑक्सीडेज (Cytochrome oxidase), एल्कोहॉल डिहाइड्रोजिनेज (Alcohol dehydrogenase), रिडक्टेज (Reductase)
– ट्रांसफेरेजेज (Transferases) – ये एन्जाइम अपने क्रियाधारों में हाइड्रोजन के अतिरिक्त किसी भी मूलक अथवा समूह को एक क्रियाधार से दूसरे में स्थानान्तरण करते हैं। इनके द्वारा स्थानान्तरित होने वाले प्रमुख समूह अमीनो, फास्फेट, मिथाइल, थायोल इत्यादि हैं।
जैसे – ट्रांसएमिनेज, ट्रांसफास्फेटेज।
– हाइड्रोलेजेज (Hydrolases) – इस वर्ग के एन्जाइम जल के अणुओं को जोड़ने अथवा निकालने वाली अभिक्रियाओं को उत्प्रेरित करते हैं। किसी भी रासायनिक अभिक्रिया में जल के अणुओं का जुड़ना हाइड्रोलेजेज एन्जाइम की उपस्थिति में होता है। ये एन्जाइम वृहत् अणुओं को जल संयोजन द्वारा छोटे-छोटे अणुओं में तोड़ देते हैं।
जैसे– एस्ट्रेज (Eseterase), कार्बोहाइड्रेज (Carbohydrase), एमाइलेज (Amylase), न्यूक्लिऐज (Nuclease)।
– लायेजेज (Lyases) – इस प्रकार के एन्जाइम अपने क्रियाधारों में से बिना जल अपघटन किए विशेष प्रकार के सह संयोजक बन्ध तोड़कर समूह निष्कासन को उत्प्रेरित करते हैं।
जैसे – एल्डोलेज (Aldolase)।
– आइसोमरेजेज (Isomerases) – इस प्रकार के एन्जाइम क्रियाधारों में अंतर आण्विक पुन:व्यवस्था के द्वारा उनकी स्थितिज अथवा प्रकाशीय समावयवी में परिवर्तन होने वाली अभिक्रियाओं को उत्प्रेरित करते हैं अर्थात् किसी क्रियाधार को उसके समावयवी अणु में परिवर्तित करते हैं।
जैसे– फास्फोहेक्सो आइसोमरेज (Phosphoexoisomerase)।
– लाइगेजेज (Ligases) – इस प्रकार के एन्जाइम दो यौगिकों को सहसंयोजक बंधों द्वारा जोड़ने वाली अभिक्रियाओं को उत्प्रेरित करते हैं।
जैसे – पायरूवेट कार्बोक्सीलेज (Pyruvate carboxylase), सिट्रेट सिन्थेटेज (Citrate synthatase)।
पादपों में श्वसन
– कार्बनिक पदार्थों का ऑक्सीकृत अपघटन (C-C बंध तोड़ना) ऊर्जा संश्लेषण के लिए कोशिकीय श्वसन कहलाता है।
– प्रोकैरियोट्स में, श्वसन कोशिका द्रव्य में होता है जबकि युकैरियोट्स में यह कोशिकाद्रव्य तथा माइटोकॉन्ड्रिया में होता है।
– कोशिकीय श्वसन – Exergonic (energy released), Amphibolic प्रक्रिया।
श्वसन के प्रकार (Types of Respiration)
– इसके दो मुख्य प्रकार हैं–
(1) ऑक्सी अथवा वायुश्वसन (Aerobic Respiration)
(2) अनॉक्सी अथवा अवायुश्वसन (Anaerobic Respiration)
1. ऑक्सी अथवा वायुश्वसन (Aerobic Respiration) – इस प्रकार का श्वसन ऑक्सीजन की उपस्थिति में अर्थात् O2 का उपभोग करते हुए सम्पन्न होता है जिसमें कार्बनिक खाद्य पदार्थ का जल एवं कार्बन डाइऑक्साइड में पूर्ण अपघटन हो जाता है एवं अधिक मात्रा में ऊर्जा मुक्त होती है।
– सभी जन्तुओं एवं पौधों में श्वसन की यह सामान्य विधि है। ऑक्सी श्वसन को निम्नलिखित रासायनिक समीकरण द्वारा दर्शाया जाता है।
C6H12O6+6O2 à 6CO2+6H2O+686k.cal ऊर्जा
2. अनॉक्सी अथवा अवायुश्वसन (Anaerobic Respiration) – इस प्रकार का श्वसन ऑक्सीजन के उपयोग के बिना होता है अर्थात् इसमें O2 का उपभोग नहीं होता है। जिससे कार्बनिक खाद्य पदार्थ का पूर्ण ऑक्सीकरण नहीं होता है एवं एल्कोहॉल अथवा कार्बनिक अम्ल तथा CO2 का निर्माण होता है। इस श्वसन में कम मात्रा में ऊर्जा मुक्त होती है। यह अंतराणुक श्वसन (Intramolecular Respiration) भी कहलाता है। अनॉक्सी श्वसन को निम्नलिखित रासायनिक अभिक्रिया द्वारा प्रदर्शित किया जाता है।
C6H12O6 à 2C2H5OH+2CO2+56k.cal ऊर्जा
– संगृहीत व अंकुरित होते हुए बीजों, मांसल फलों में अस्थायी रूप से एवं कई कवकों तथा जीवाणुओं में नियमित रूप से अनॉक्सी श्वसन पाया जाता है।

ऑक्सी व अनॉक्सी श्वसन में सम्बन्ध
ऑक्सी श्वसन व अनॉक्सी श्वसन में अन्तर –
| क्र. सं. | ऑक्सी श्वसन | अनॉक्सी श्वसन |
| 1. | इसमें O2 का उपभोग होता है। | इसमें O2 की आवश्यकता नहीं होती है। |
| 2. | सभी जीवित कोशिकाओं में होता है। | यह केवल कुछ कवकों, जीवाणुओं, कोशिकाओं तथा संगृहीत एवं अंकुरित होते हुए बीजों आदि में होता है। |
| 3. | इस श्वसन में ग्लूकोज के एक अणु के पूर्ण ऑक्सीकरण के द्वारा 38 ATP अणुओं का निर्माण होता है। | इसमें ग्लूकोज का पूर्ण ऑक्सीकरण नहीं होता है तथा केवल 2ATP का निर्माण होता है। |
| 4. | इस श्वसन में होने वाली ग्लाइकोलिसिस, कोशिका द्रव्य में तथा क्रेब्स चक्र माइटोकॉन्ड्रिया में सम्पन्न होती है। | सभी क्रियाएँ कोशिकाद्रव्य में सम्पन्न होती हैं। |
| 5. | इसमें श्वसन पदार्थ का पूर्ण ऑक्सीकरण होता है। | इसमें श्वसनी पदार्थ का अपूर्ण ऑक्सीकरण होता है। |
| 6. | अन्त्योत्पाद CO2, जल एवं ऊर्जा होते हैं। | अन्त्योत्पाद एल्कोहॉल अथवा कार्बनिक अम्ल, CO2, एवं ऊर्जा होते हैं। |
– श्वसन क्रियाधार/श्वसनाधार (Respiratory Substrate) – जीवित कोशिकाओं में वे कार्बनिक यौगिक जो केटाबॉलिज्म द्वारा ऊर्जा मुक्त करते हैं, उन्हें श्वसन धार कहते हैं। उदाहरण – कार्बोहाइड्रेट, वसाएँ, प्रोटीन्स, कार्बनिक अम्ल। ब्लैकमैन ने श्वसन धार के आधार पर इसे अनेक नाम दिए। यदि श्वसन में कार्बोहाइड्रेट्स या वसाएँ प्रयुक्त हों तो इसे ‘प्लावी श्वसन’ (Floating respiration) कहते हैं जबकि प्रोटीन प्रयुक्त होने पर इसे जीवद्रव्यी श्वसन (Protoplasmic respiration) कहते हैं।
वायवीय श्वसन की क्रियाविधि (Mechanism of Aerobic Respiration)–
– इसमें दो प्रकार शामिल हैं – कॉमन पाथवे तथा पेन्टोज फॉस्फेट पाथवे।
1. वायवीय श्वसन का कॉमन पाथवे (Common Pathway of Aerobic Respiration) – ग्लाइकोलिसिस वायवीय श्वसन तथा अवायवीय श्वसन में उभयनिष्ठ होता है इसलिए इसे कॉमन पाथवे कहते हैं। वायवीय श्वसन के कॉमन पाथवे का प्रक्रम निम्नलिखित चरणों में पूर्ण होता है–
1. ग्लाइकोलिसिस (Glycolysis) – यह आंशिक ऑक्सीकारी प्रक्रिया है जिसमें हेक्सोज (ग्लूकोज या फ्रक्टोज) पायरुविक अम्ल के दो अणुओं में टूट जाता है। इसे EMP परिपथ भी कहते हैं क्योंकि इसकी खोज तीन जर्मन वैज्ञानिकों एम्बडन, मेयरहॉफ तथा पारनास ने की थी।

ग्लाइकोलिसिस में ऊर्जा उत्पादन
(Energy Production in Glycolysis) –
– ग्लाइकोलिसिस में हेक्सोज शर्करा के एक अणु के टूटने से 3-C यौगिक पायरुविक अम्ल के दो अणु बनते हैं। ATP के चार अणु बनते हैं तथा ATP के 2 अणु प्रयुक्त होते हैं अतः शुद्ध लाभ 2 ATP का होता है।
– 2 अणु NADPH.H+ के भी बनते हैं जो माइटोकॉन्ड्रिया में प्रवेश करते हैं तथा ETS द्वारा ऑक्सीकृत होकर 6ATP बनाते हैं। इस तरह वायुवीय श्वसन में ग्लाइकोलिसिस से 2+6 = 8ATP बनते हैं।
2. पायरुविक अम्ल का ऑक्सीकारी विकार्बोक्सिलीकरण (Oxidative decarboxylation of pyruvic acid) –
– पायरुविक अम्ल (3C) का ऑक्सीकारी विकार्बोक्सिलीकरण होता है तथा एसीटिल Co-A बनता है। यह माइटोकॉन्ड्रिया में होता है।
– यह अभिक्रिया एन्जाइम पायरुवेट डिहाइड्रोजिनेज तथा TPP.CoA, लिपोइक अम्ल, NAD+ तथा Mg++ आयन्स द्वारा उत्प्रेरित होती है।

3. क्रेब्स चक्र (Krebs cycle) –
– इसे ट्राइकार्बोक्सिलिक अम्ल चक्र (TCA चक्र) या सिट्रिक अम्ल चक्र (CA चक्र) भी कहते हैं क्योंकि इसके प्रथम स्थायी उत्पाद सिट्रिक अम्ल में तीन कार्बोक्सिलिक समूह (-COOH) होते हैं।
– यह माइट्रोकॉन्ड्रिया के मैट्रिक्स भाग में होता है।
– एसीटिल Co A ग्लाइकोलिसिस तथा क्रेब्स चक्र के बीच की योजक कड़ी है।
– इसमें एनाबॉलिक तथा केटाबॉलिक अभिक्रियाएँ दोनों शामिल हैं इस प्रकार यह एम्फीबॉलिक चक्र है। क्रेब्स चक्र में एसीटिल CoA (20) ओक्जेलो एसीटिक अम्ल (4C) के साथ क्रिया कर सिट्रिक अम्ल (6C) बनाता है।

क्रेब्स चक्र
– एक क्रेब्स चक्र द्वारा एसीटिल CoAका एक अणु 3 NADH.H+, 1 FADH.H+ तथा 1 ATP (=12 ATP) बनाता है।
– इस प्रकार पायरुविक अम्ल का एक अणु 4NADH.H+, 1 FADH.H+, तथा 1 ATP अणु (= 15ATP) उत्पन्न करता है। इस तरह पायरुविक अम्ल के दो अणु 30 ATP बनाएँगे। इस प्रकार पायरुविक अम्ल का एक अणु क्रेब्स चक्र द्वारा होने वाले ऑक्सीकरण से 3 अणु CO2 बनाता है। इस प्रकार पायरुविक अम्ल के दो अणु CO2 के 6 अणु उत्पन्न करते हैं।
4. टर्मीनल ऑक्सीडेशन (Terminal oxidation) –
– केटाबॉलिक प्रक्रम के अन्त में वायुमण्डलीय O2 शामिल होती है। इसमें दो प्रक्रम शामिल है।
(A) इलेक्ट्रॉन परिवहन तंत्र या (ETS)
(B) ऑक्सीकरण फॉस्फोरिलीकरण
(A) इलेक्ट्रॉन परिवहन तंत्र या ETS (Electron Transport System or ETS)–
– यह F1 कण या फर्नांडिस मोरान उपइकाइयों या ऑक्सीसोम्स में होता है जो माइटोकॉन्ड्रिया की आन्तरिक कला पर स्थित होते हैं।
– प्रत्येक समूह में एन्जाइम्स एक विशिष्ट श्रेणी में व्यवस्थित होते हैं। जिसे इलेक्ट्रॉन परिवहन शृंखला (Electron transport chain) या माइटोकॉन्ड्रियल श्वसनीय शृंखला या इलेक्ट्रॉन परिवहन तंत्र (Electron transport system or ETS) कहते हैं।
प्रक्रम (Process) – माइट्रोकॉन्ड्रिया की आन्तरिक कला में पाँच कॉम्प्लेक्स (Complexes) होते हैं।
| S. No. | Name of Complexes | Parts of ETS |
| 1. | Complex-I | FMN-NADH2 Dehydrogenase |
| 2. | Complex-II | CoQ/UQ-FADH2 Dehydrogenase/ Succinate dehydrogenase |
| 3. | Complex-III | Cyt. b-Cytc1 |
| 4. | Complex-IV | Cyt. a & Cyt.a3 (Cu Present) |
| 5. | Complex-V | ATP synthase/ATPase |
– कॉम्प्लेक्स I से IV इलेक्ट्रॉन परिवहन में शामिल होते हैं जबकि कॉम्प्लेक्स V (F0-F1 कण) ATP संश्लेषण से संबंधित होता है। इसमें निम्नलिखित चरण शामिल हैं–

इलेक्ट्रॉन परिवहन तंत्र
(i) इलेक्ट्रॉन परिवहन तंत्र (ETS) फ्लेविन्स, Fes कॉम्प्लेक्स, क्विनॉन तथा सायटोक्रोम्स का बना होता है। सामान्य क्विानॉन यूर्बीक्विनॉन या CoQ है। इलेक्ट्रॉन्स माइटोकॉन्ड्रियल इलेक्ट्रॉन परिवहन शृंखला में दो पथों से प्रवेश करते हैं। FMN पर तथा FAD’ पर।
(ii) NADH अपने इलेक्ट्रॉन तथा H+ आयन्स FMN को स्थानान्तरित करते हैं जो कि ETC के पथ 1 में प्रथम इलेक्ट्रॉन ग्राही है। इस स्थानान्तरण में NADH ऑक्सीकृत होता है तथा FMN, FMNH2 में अपचयित हो जाता है।
क्रेब्स चक्र में सक्सीनेट से मुक्त इलेक्ट्रॉन्स FAD+ द्वारा ग्रहण किए जाते हैं। जो कि ETC के पथ 2 में प्रथम इलेक्ट्रॉन वाहक हैं।
(iii) H- आयन माइटोकॉन्ड्रिया के बाह्य कक्ष में गति करते हैं जबकि इलेक्ट्रॉन्स साइटोक्रोम्स द्वारा ग्रहण किए जाते हैं। साइटोक्रोम्स बढ़ते हुए रेडॉक्स विभव के अनुसार व्यवस्थित होते हैं।
Cyt b→Cyt C1→Cyt C→Cyt a→cyt a3
साइटोक्रोम a तथा a3 को संयुक्त रूप से साइटोक्रोम ऑक्सीडेज कहते हैं जबकि साइटोक्रोम a3 को टर्मीनल ऑक्सीडेज कहते हैं।
(iv) साइटोक्रोम में, आयरन सक्रियकारक के रूप में कार्य करता है। यह इलेक्ट्रॉन को ग्रहण करने (Fe+++ + e– Fe++) तथा देने (Fe++ – e– → Fe+++) का कार्य करता है।
(v) साइटोक्रोम ऑक्सीडेज से ये इलेक्ट्रॉन निकलकर ऑक्सीजन के परमाणु से क्रिया कर उसे सक्रिय बनाते हैं। यह सक्रिय ऑक्सीजन 2 हाइड्रोजन आयनों के साथ क्रिया कर जल का अणु बनाता है।

(B) ऑक्सीकारी फॉस्फोरिलीकरण
(Oxidative Phosphorylation) –
– माइटोकॉन्ड्रिया में ऑक्सीजन की उपस्थिति में ATP का संश्लेषण ऑक्सीकारी फॉस्फोरिलीकरण कहलाता है। जो कि रसोपरासरणी सिद्धान्त (Chemiosmatic theory) द्वारा समझाया जा सकता है।
रसोपरासरणी सिद्धान्त (Chemiosmotic theory) –
– इसका प्रतिपादन पीटर मिशैल (1961) ने किया, उन्हें 1978 में नोबेल पुरस्कार मिला। इस सिद्धान्त के अनुसार NADH के ऑक्सीकरण के दौरान H+ आयन्स माइटोकॉन्ड्रिया के बाह्य कक्ष में चले जाते हैं। जिसके कारण कला के आर-पार pH में अन्तर उत्पन्न होता है जो माइटोकॉन्ड्रिया की आन्तरिक कला के आर-पार विद्युत रासायनिक विभव उत्पन्न करती है जो प्रोटोन प्रवणता या प्रोटोन गतिदायी बल उत्पन्न करता है।
– प्रोटोन गतिदायी बल प्रोटोन्स के बाह्य कक्ष से आन्तरिक कला को पार कर मैट्रिक्स में प्रवाह को प्रेरित करता है। प्रोटोन F1 कण से गुजरते हैं जहाँ ATPase ADP तथा Pi से ATP निर्माण को प्रेरित करता है।
ETS का पथ -1 : एक अणु NADH के पूर्ण ऑक्सीकरण द्वारा
3ATP का संश्लेषण होता है।
ETS का पथ -2 : एक अणु FADH के पूर्ण
ऑक्सीकरण द्वारा 2 ATP का संश्लेषण होता है।
2. श्वसन का पेन्टोज फॉस्फेट पाथवे (Pentose phosphate pathway of respiration)–
– इस चक्र मे ग्लूकोज 6-P के छ: अणु प्रवेश करते हैं। ऑक्सीकरण के बाद ये CO2 के 6 अणु तथा कोएन्जाइम NADPH2 के 12 अणु उत्पन्न करते हैं जबकि 5 अणु ग्लूकोज 6 – P पुनरुत्पादित होते हैं।
– 12 NADPH+H+ ETS मे ऑक्सीकृत हो कर 36ATP बनाते हैं लेकिन ग्लूकोज के फॉस्फोरिलीकरण में 1ATP का उपभोग होता है। इसलिए ATP का शुद्ध लाभ 35 ATP होता है।
– HMP या PPP के मुख्य चरण निम्नलिखित है। (HMP-हेक्सोज मोनोफॉस्फेट पाथवे)
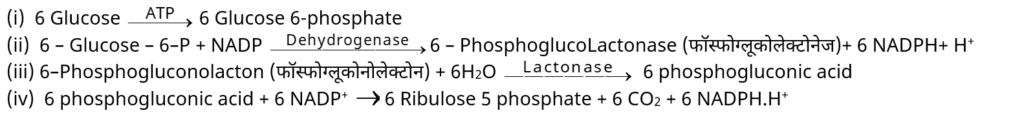
– राइबुलोज – 5- P के 6 अणु ग्लूकोज -6-P के 5 अणु बनाते हैं।
अनॉक्सी अथवा अवायुश्वसन (Anaerobic Respiration) – इसमें आण्विक ऑक्सीजन प्रयुक्त नहीं होती है तथा CO2 के निर्माण सहित या उसके बिना कार्बनिक भोज्य पदार्थ अपूर्ण रूप से ऑक्सीकृत होते हैं। यह कम मात्रा मे ऊर्जा मुक्त करते हैं। इसे अन्तः आण्विक श्वसन (intramolecular respiration) भी कहते हैं।

-Oxidation of Fatty acids
Glyoxisome, peroxisome cytosol
Fatty acids, 2,carbon, Acetyl CoA, ATP
𝛽-oxidation

किण्वन (Fermentation) –
– इसमें सूक्ष्मजीवियों की सहायता से कार्बोहाइड्रेटस तथा अन्य कार्बनिक यौगिकों के अवायवीय विघटन से एल्कोहॉल, कार्बनिक अम्ल तथा CO2 आदि का निर्माण होता है यह ऑक्सीजन की अनुपस्थिति में होता है।
अवायवीय श्वसन की क्रियाविधि (Mechanism of Anaerobic Respiration) – इसमें दो चरण शामिल हैं–
(i) ग्लाइकोलिसिस (Glycolysis) – 2NADH. H+ की अनुपस्थिति को छोड़कर यह वायवीय श्वसन के ग्लाइकोलिसिस के समान होता है।
(ii) पायरुविक अम्ल का अवायवीय विघटन (Anaerobic breakdown of pyruvic acid) – जीव, ऊतक के प्रकार तथा अन्तिम उत्पाद की प्रकृत्ति के आधार पर अवायवीय श्वसन या किण्वन में निम्नलिखित प्रकार शामिल हैं।
1. एल्कोहॉलिक किण्वन (Alcoholic fermentation) – यह प्रक्रम ग्लूकोज से पायरुविक अम्ल के निर्माण द्वारा प्रारम्भ होता है। पायरुविक अम्ल द्वारा एल्कोहॉल के निर्माण के दो चरण हैं।
यह जीवाणु, कवक तथा उच्च पादपों में होती है।
2. लेक्टिक अम्ल किण्वन (Lacticacid fermentation) – कुछ जीवाणु पायरुविक अम्ल को लेक्टिक अम्ल में परिवर्तित कर देते हैं। जैसे – लेक्टोबैसीलस लेक्टिस।

श्वसनीय गुणांक या साँस गुणांक (Respiratory Quotient or R.Q.) –
– श्वसन में मुक्त CO2 तथा ग्रहण की गई ऑक्सीजन के आयतन का अनुपात R.Q. है।

– RQ का मापन गेनांग रेस्पिरोमीटर द्वारा किया जा सकता है।
1. कार्बोहाइड्रेट्स का R.Q. :
– यदि कार्बोहाइड्रेटस श्वसनाधार है तब R.Q. एक होता है।
C6H12O6+6O2 →6CO2 + 6H2O + 686 K. cals.
𝑅𝑄=6𝐶𝑂26𝑂2=66=1
2. वसाओं का R.Q. –
– जब वसाएँ श्वसनीय पदार्थ हैं तो R.Q. का मान 1 से कम हो जाता है। वसाओं में ऑक्सीजन कम होती है तथा इन्हें ऑक्सीकृत होने के लिए अधिक O2 की आवश्यकता होती है।
2C51H98O6 + 145O2 (Tripalmatin) →102CO2 + 98H2O + E
C57H104O6 (Triolin) + 8002 → 57CO2 + 52H2O + E
R.Q = 57𝐶𝑂280𝑂2=5780=0.71
3. प्रोटीन्स का R.Q. –
– जब प्रोटीन्स श्वसनाधार हैं तो R.Q. का मान एक से कम (सामान्यतः 0.9) होता है।
4. कार्बनिक अम्ल का R.Q. –
– जब श्वसन में कार्बनिक अम्ल (प्रकाश की उपस्थिति में गूद्देदार पादपों में) ऑक्सीकरण होते हैं तो R.Q. का मान 1 से अधिक होता है, क्योंकि कार्बनिक अम्लों में O2 प्रचुर होती है तथा उन्हें ऑक्सीकरण के लिए कम O2 चाहिए।
2(COOH)2 +O2→ 4CO2+2H2O+E Oxalic acid
𝑅.𝑄=4𝐶𝑂2𝑂2=41=4
C4H6O5 + 3O2 (मैलिक अम्ल) → 4CO2 + 3H2O + E
𝑅.𝑄=4𝐶𝑂23𝑂2=43=1.33
– मांसल पादपों या सरस पादपों का R.Q शून्य होता है।
श्वसन को प्रभावित करने वाले कारक (Factors Affecting Respiration) –
1. तापक्रम 6. खनिजलवण
2. प्रकाश 7. श्वसनाधार
3. CO2 8. कार्बनिक पदार्थ
4. O2 9. आयु
5. जल 10. जीवद्रव्यी कारक
1. तापक्रम – अनुकूलतम तापमान = 30°C
Q10 मान = 2–2.5
0°C से कम ताप पर श्वसन की दर बहुत तेजी से घटती है। यद्यपि कुछ पौधों में श्वसन -20°C पर भी होता है। प्रसुप्त बीज -50°C पर भी जीवित रहते हैं।
2. प्रकाश – प्रकाश श्वसन की दर को अप्रत्यक्ष रूप से प्रभावित करता है।
प्रकाश में वृद्धि → प्रकाश संश्लेषण की दर में वृद्धि → श्वसन के लिए पदार्थों में वृद्धि → श्वसन की दर में वृद्धि
3. CO2 – यदि CO2 की मात्रा वायु में सामान्य से अधिक है तो श्वसन की दर घटती है, बीज का अंकुरण घटता है तथा वृद्धि भी घटती है।
4. O2 – वातावरण में ऑक्सीजन की मात्रा में अल्पवृद्धि या कमी से श्वसन की दर में कोई प्रभाव नहीं पड़ता है।
वातावरण में ऑक्सीजन की मात्रा 1.9% तक घटाने पर ऑक्सीश्वसन नगण्य हो जाता है, इसे ऑक्सीश्वसन का विलोपन बिन्दु कहते हैं लेकिन अनॉक्सी श्वसन होता है।
5. जल – श्वसन की दर ∝ जल की मात्रा
सूखे बीजों में श्वसन की दर बहुत कम होती है लेकिन जैसे ही वे जल का अंतःशोषण करते हैं, श्वसन की दर बढ़ जाती है।
6. खनिज लवण – क्षारीय मृदा तत्त्वों के क्लोराइड तथा डाइवैलेन्ट कैटायन (Li, Ca, Mg) श्वसन की दर को बढ़ा देते हैं।
7. श्वसन के लिए उपलब्ध पदार्थ – श्वसन की दर ∝ श्वसन के लिए उपलब्ध पदार्थ
8. प्रदूषक – गैसीय प्रदूषक जैसे SO2 NO2 तथा O3 की अधिक सांद्रता कला की क्षति करती है जिससे श्वसन की दर घट जाती है। ये गैसीय प्रदूषक pH बढ़ाते हैं जो कि ETS को प्रभावित करता है।
9. आयु – आयु तथा परिपक्वता के साथ श्वसन की दर बढ़ती है।
10. जीव द्रव्यी कारक – अधिक जीवद्रव्य → अधिक श्वसन की दर
विभज्योत्की कोशिकाओं में श्वसन की दर परिपक्व कोशिकाओं की अपेक्षा अधिक होती है।
प्रकाश संश्लेषण
– CO2 तथा H2O की सहायता से प्रकाश की उपस्थिति में पादपों के हरे भागों द्वारा जटिल कार्बनिक पदार्थों (कार्बोहाइड्रेट्स) का संश्लेषण प्रकाश संश्लेषण कहलाता है।

– प्रकाश संश्लेषण एनाबॉलिक, एण्डोथर्मिक तथा रेडॉक्स प्रक्रम है।
– प्रकाश संश्लेषण सबसे पहले सायनोबैक्टीरिया में देखा गया।
– प्रकाश संश्लेषण की प्रक्रिया में सूर्य की विकिरण ऊर्जा रासायानिक ऊर्जा में परिवर्तित हो जाती है। इसे कार्बन डाई ऑक्साइड स्वांगीकरण (Carbon dioxide assimilation) भी कहते हैं।
– प्रकाश संश्लेषण एक रेडॉक्स प्रक्रिया है जिसमें जल के ऑक्सीकरण से O2 बनती है जबकि CO2 अपचयित होकर शर्कराएँ बनाती है इस प्रकार यह ऑक्सीकरण अपचयन-अभिक्रिया है।
प्रकाश संश्लेषण के स्थल (Site of photosynthesis) –
– प्रकाश संश्लेषण की क्रिया क्लोरोप्लास्ट में होती है।
– पादपों की Mesophyill कोशिकाओं के क्लोरोप्लास्ट में होती है।
– प्रत्येक थाइलेकॉइड लिपिड तथा प्रोटीन की बनी एक कला होती है। वे सिक्के के चट्टे की भाँति व्यवस्थित होती है। इन्हें ग्रेना (एकल ग्रेनम) कहते हैं।
– दो ग्रेना एक लम्बी पटलिका (Lamellae) द्वारा अन्तर्सम्बन्धित होती हैं। जिसे इन्टरप्रेनम या फ्रेट चैनल कहते हैं।
– अनेक छोटे कण थाइलेकॉइड कला पर पाए जाते हैं। जिन्हें क्वाण्टोसोम्स कहते हैं इनकी खोज पार्क तथा विगिन्स ने की।
वर्णक (Pigments) –
– क्लोरोप्लास्ट में वर्णक पाए जाते हैं।
1. क्लोरोफिल 2. जैन्थोफिल 3. कैरोटिन

क्लोरोफिल (Chlorophyll) –
– क्लोरोफिल की संरचना (Structure of chlorophyll) –
– क्लोरोफिल का अणु टेडपोल सदृश्य होता है।
– प्रत्येक क्लोरोफिल अणु दो भागों का बना होता है। पोरफाइरिन शीर्ष तथा फाइटॉल पुच्छ। शीर्ष टेट्रापायरोल या पोरफाइरिन का बना होता है जिसमें चार पायरोल अणु समचक्रीय रूप से व्यवस्थित होते हैं। जिनके केन्द्र में एक Mg होता है।
– पायरोल अणु में कार्बन तथा नाइट्रोजन का ढाँचा होता है जिसमें वलय संरचना होती है। इनमें से एक पायरोल वलय लम्बी शृंखला वाले एल्कोहॉल से जुड़कर इसकी पुच्छ बनाते हैं।
– क्लोरोफिल a में मिथाइल समूह (–CH3) होता है तथा क्लोरोफिल b में एल्डिहाइड समूह (–CHO) होता है। chl b, chl a का एल्डिहाइड रूप है।
chl – a – C55H72O5N4 Mg
chl – b – C55H70O6N4 Mg
– केरोटिनॉइड्स (Carotenoids) –
– अधिकांश केरोटिनॉइड्स पीले या नारंगी रंग के होते हैं। लिपिड्स में घुलनशील होते हैं। वे क्लोरोफिल को प्रकाश ऑक्सीकरण से बचाते हैं। इस प्रकार इन्हें प्रोटेक्टिव वर्णक या शील्ड वर्णक कहते हैं।
(i) कैरोटीन (Carotene) –
– ये नारंगी रंग के होते हैं। ये हाइड्रोकार्बन्स हैं जिनका सामान्य सूत्र C40H56 होता है।
– कैरोटीन के तीन मुख्य समावयव हैं – 𝛼– कैरोटीन, 𝛽– कैरोटीन, 𝛾– कैरोटीन।
– लाइकोपिन (C40H51) कैरोटीन है जो टमाटर में पाया जाता है।
(ii) जेन्थोफिल (Xanthophyll) –
– इन्हें कैरोटिनॉल्स भी कहते हैं। ये भूरे रंग के होते हैं। ये कैरोटीन के ऑक्सीकृत व्युत्पन्न हैं।
उदाहरण – C40H56O2 – ल्यूटीन
– अन्य महत्त्वपूर्ण जेन्थोफिल्स हैं। क्रिप्टोजैन्थिन, वायलोजैन्थिन, फ्लेवोजैन्थिन। फ्यूकोजेन्थिन भूरे शैवाल में पाया जाता है।
अवशोषण तथा क्रिया स्पेक्ट्रा
(Absorption and action spectra)
अवशोषण स्पेक्ट्रम
– दृश्य प्रकाश की तरंगदैर्ध्य की परास 3900 𝐴𝑜 – 7600 𝐴𝑜 होती है। दृश्य प्रकाश के कणों को फोटोन कहते हैं।
– दृश्य प्रकाश के सात रंगों (VIBGYOR) में से हरा प्रकाश अवशोषित नहीं होता है पुनः परावर्तित हो जाता है तथा पत्तियों को हरा रंग प्रदान करता है।
– Chl a अधिकतम अवशोषण नीले तथा लाल प्रकाश का करता है। एक पदार्थ द्वारा अवशोषित विभिन्न तरंगदैर्ध्यों के प्रकाश की मात्रा को दर्शाने वाला वक्र अवशोषण स्पेक्ट्रम कहलाता है।
क्रिया स्पेक्ट्रम (Action spectrum) –
– यह एक ग्राफ है जो प्रकाश की विभिन्न तंरगदैर्ध्यों पर O2 उत्पादन के सम्बन्ध में मापी गई प्रकाश संश्लेषण की दर को दर्शाता है। यह लाल में अधिकतम फिर नीले में तथा सबसे कम हरे प्रकाश में होता है।
– इन्जेलमैन ने प्रिज्म की सहायता से प्रकाश को इसके अलग-अलग रंगों में विभक्त किया और फिर उसे हरी शैवाल, क्लेडोफोरा को दिया जिसे बैक्टीरिया के साथ रखा गया। बैक्टीरिया का उपयोग O2 निष्कासन का स्थान ज्ञात करने के लिए किया गया था। उन्होंने देखा कि बैक्टीरिया मुख्यतः प्रकाश के लाल और नीले हिस्से में एकत्रित हो जाते हैं। इस प्रकार सबसे पहले, क्रिया स्पेक्ट्रम का विवरण दिया गया। ये लगभग क्लोरोफिल a तथा b के अवशोषण स्पेक्ट्रम से समानता दर्शाता है।
लाल पतन (Red drop) तथा इमरसन संवृद्धि प्रभाव (Emerson enhancement effect) –
– इमरसन ने जब विभिन्न तरंगदैर्ध्यों के एकवर्णीय प्रकाश के उपयोग द्वारा क्लोरेला में प्रकाश संश्लेषण की क्वाण्टम लब्धि को निर्धारित किया तो उन्होंने देखा कि 680nm से अधिक तंरगदैर्ध्य पर क्वाण्टम लब्धि तेजी से कम हो जाती है। प्रकाशसंश्लेषी उत्पादन में कमी चूँकि स्पैक्ट्रम के लाल क्षेत्रों से सम्बन्धित है तो इसे लाल पतन कहते हैं।
– इमरसन ने अवरक्त प्रकाश (660 nm. से ज्यादा) के साथ-साथ इससे छोटी तरंगदैर्ध्य का प्रकाश भी दिया तो उसने पाया कि क्वाण्टम लब्धि में बढ़ोतरी हुई। इसे इमरसन संवृद्धि प्रभाव कहते हैं।
– इमरसन ने इस प्रयोग के बाद दो वर्णक तंत्रों (Pigment systems) की उपस्थिति को स्पष्ट किया।
दो वर्णक तंत्र (Two pigment system)
(i) वर्णक तंत्र (ii) वर्णक तंत्र-II
1. PS-I या वर्णक तंत्र-I (LHC I या Light harvesting complex-I) –
– यह ग्रेना थाइलेकॉइड्स व स्ट्रोमा थाइलेकॉइड्स दोनों के प्रभाव रहित भाग पर स्थित होता है।
– यह लम्बी तरंगदैर्ध्य वाले प्रकाश को अवशोषित करने वाले वर्णकों का बना होता है। chla -P 700 (अभिक्रिया केन्द्र), chl b, कैरोटिनॉइड्स, साइटोक्रोम कॉम्प्लेक्स, प्लॉस्टोसायनिन तथा फेरीडॉक्सिन होते हैं।
2. PS-II या वर्णक तंत्र-II (LHC II या Light harvesting complex-II) –
– यह ग्रेना थाइलेकॉइड के सटे हुए भाग पर स्थित होता है।
– यह छोटी तंरगदैर्ध्य वाले प्रकाश को अवशोषित करने वाले वर्णक का बना होता है। chla – 680 (अभिक्रिया केन्द्र), कैरोटिनॉइड्स, प्लास्टोक्विनॉन, मैंगनीज तथा क्लोराइड, क्वेन्चर अणु Q, साइटोक्रोम जटिल तथा प्लास्टोसायनिन युक्त होता है।
रसायनी संश्लेषण (Chemosynthesis)
– कुछ वायवीय जीवाणु अकार्बनिक पदार्थों के ऑक्सीकरण द्वारा मुक्त होने वाली रासायनिक ऊर्जा का उपयोग कार्बनिक (भोज्य) पदार्थ बनाने में करते हैं, यह क्रिया रासायनी संश्लेषण (Chemosynthesis) कहलाती है।
कुछ महत्त्वपूर्ण रासायनी संश्लेषण करने वाले जीवाणु निम्नलिखित हैं–
– नाइट्रोसोमोनास व नाइट्रोकोकस – ये जीवाणु अमोनिया का ऑक्सीकरण नाइट्राइट में करते हैं व इस क्रिया में मुक्त होने वाली रासायनिक ऊर्जा का उपयोग कार्बनिक पदार्थ बनाने में किया जाता है।
– आयरन जीवाणु – जैसे फैरोबेसिलस व लेप्टोथ्रिक्स आदि फेरस (Fe+2) का ऑक्सीकरण फैरिक (Fe+3) में करते हैं व मुक्त होने वाली ऊर्जा का उपयोग कार्बनिक पदार्थों के संश्लेषण में करते हैं।
– रसायनी संश्लेषण के कुछ अन्य उदाहरण सल्फर जीवाणु (थायोथ्रिक्स), हाइड्रोजन जीवाणु (बेसिलस पेन्टोट्रोपस) व कार्बन जीवाणु (बेसिलस ओलिगोकार्बोफिलस) हैं।
2NH3 + 3O2→2HNO2 + 2H2O + ऊर्जा ↑
– कार्बन जीवाणु में बेसिलस ओलिगोकार्बोफिलस (Bacillus oligocarbophilus)।
2CO +O2 → 2CO2 + ऊर्जा↑
प्रकाश-संश्लेषण की क्रियाविधि
1. Light reaction (प्रकाशिक अभिक्रिया)/Hill reaction (हिल अभिक्रिया) Photophosphorylation (प्रकाश फॉस्फोरीलीकरण अभिक्रिया)/Production of Assimilatory powers (स्वांगीकरण शक्ति NADPH2 & ATP का निर्माण।)
– प्रकाशिक अभिक्रिया के दौरान दोनों वर्णक तंत्रों की प्रतिक्रियाओं द्वारा प्रकाशीय ऊर्जा को रासायनिक ऊर्जा (ATP) में परिवर्तित किया जाता है। ये क्रियाएँ चक्रीय, अचक्रीय प्रकार की होती हैं।
– एंटीना वर्णक सहायक वर्णक विकिरण ऊर्जा प्राप्त करते हैं व इसे एक-दूसरे पर स्थानांतरित करते हैं। इस घटना को अनुनाद स्थानांतरण (Resonance transfer) कहते हैं। इसी प्रकार एंटीना वर्णक उत्तेजित होकर अपनी ऊर्जा अभिक्रिया केन्द्र के Chl. a को देते हैं।
(I) Cyclic ETS (चक्रीय इलेक्ट्रॉन अभिगमन तंत्र) and Photophosphorylation (प्रकाश फॉस्फोरिलीकरण) – चक्रीय इलेक्ट्रॉन अभिगमन तंत्र व प्रकाश फॉस्फोरिलीकरण

– इस प्रक्रम में केवल Pigment system-I ही कार्य करता है, जो 680 nm से अधिक तरंगदैर्ध्य पर सक्रिय होता है।
– PS-I का प्रतिक्रिया केन्द्र Chl.-a- 700/P-700 होता है जिसका e– उत्तेजित होकर ETS में चलता है।
– चक्रीय ETS के दौरान ऑक्सीजन नहीं निकलती क्योंकि इसमें जल का प्रकाशिक अपघटन नहीं होता क्योंकि चक्रीय ETS के दौरान वर्णक तन्त्र से उत्तेजित इलेक्ट्रॉन वापस अभिक्रिया केन्द्र पर आ जाता है। चक्रीय ETS में दो स्थानों पर ATP बनते हैं।
– चक्रीय प्रक्रम में NADPH2 (Reducing power) का निर्माण नहीं होता है।
– चक्रीय ETS में प्लास्टोसाइनिन एकमात्र ताम्र (Cu) युक्त नीली क्रोमो प्रोटीन है।
– Cyclic ETS में e– का प्रथम ग्राही फेरेडॉक्सिन (fd) होता है। वर्तमान में Ist e– acceptor FRS (feredoxin reducing substance) माना जाने लगा है जो एक Fe-S प्रोटीन है।
(II) Non-cyclic ETS (अचक्रीय इलेक्ट्रॉन अभिगमन तन्त्र)/Z-Scheme तथा Photophosphorylation

– इस क्रिया में PS-I तथा PS-II दोनों साथ-साथ कार्य करते हैं।
– Z-स्कीम में उत्तेजित e– वापस उसी स्थान (PS-II) पर नहीं लौटता अतः e– की कमी जल के e– पूरा करते हैं। इसी क्रिया में जल के प्रकाशिक अपघटन से ऑक्सीजन गैस निकलती है तथा NADPH2 व ATP का निर्माण होता है।
– एक बार अचक्रीय फोटोफॉस्फोरिलीकरण या Z-स्कीम होने पर 2NADPH2 व 1ATP का निर्माण होता है एवं जल के चार अणुओं के प्रकाशिक अपघटन (Photolysis) होने से एक अणु ऑक्सीजन गैस निष्कासित होती है।
– Z-scheme में PS-II के उत्तेजित e– सर्वप्रथम ग्राही P.Q. होता है।
– Z-scheme के दौरान PS-I व PS-II के बीच कड़ी (PC) प्लास्टोसायनिन को माना जाता है।
– Non-cyclic ETS के अंतर्गत e– ऊर्जा का प्रवाह PS-II से PS-I की ओर होता है।
प्रकाश फॉस्फोरिलीकरण (Photophosphorylation) – प्रकाश ऊर्जा के उपयोग द्वारा ADP तथा अकार्बनिक फॉस्फेट से रासायनिक ऊर्जा या ATP के निर्माण की प्रक्रिया को प्रकाश फॉस्फोरिलीकरण कहते हैं।
अप्रकाशिक अभिक्रिया (Dark Reaction) –
– यह तापसंवेदी क्रिया है जो क्लोरोप्लॉस्ट के स्ट्रोमा में होती है।
– इस प्रक्रिया में ATP तथा NADPH का उपयोग होता है तथा CO2 के अपचयन तथा स्वांगीकरण द्वारा कार्बोहाइड्रेट्स बनते हैं। अप्रकाशिक अभिक्रिया के लिए Q10 का मान 2 या 3 होता है।
C3 चक्र या केल्विन चक्र (C3 cycle or calvin cycle) –
– अप्रकाशिक अभिक्रिया के कार्बन पथ की खोज केल्विन तथा बेन्सन (1961) ने की। उन्होंने अपने प्रयोग मे रेडियोएक्टिव C14, एक कोशिका शैवाल सिनडेस्मस, क्लोरेला का उपयोग किया।
– केल्विन चक्र को C3 चक्र भी कहते हैं क्योंकि C3 पादपों में प्रथम स्थायी यौगिक 3 कार्बन यौगिक 3- फॉस्फोग्लिसरिक अम्ल या 3-PGA अम्ल हैं।
– पादपों जिनमें C3 चक्र होता है। (अधिकांशत: समोद्भिद तथा जलोद्भिद) उन्हें C3 पादप कहते हैं।

– इसमें तीन चरण शामिल हैं–
1. कार्बोक्सिलीकरण (Carboxylation) – CO2 के यौगिकीकरण से एक स्थिर कार्बनिक मध्यस्थ बनता है। केल्विन चक्र में कार्बोक्सिलीकरण एक अत्यधिक निर्णायक चरण है जहाँ RuBP के कार्बोक्सिलीकरण के लिए CO2 का उपयोग किया जाता है। यह प्रतिक्रिया एंजाइम RuBP कार्बोक्सिलेस के द्वारा उत्प्रेरित होती है, जिसके परिणामस्वरूप 3-PGA के दो अणु बनते हैं। चूँकि इस एंजाइम में एक ऑक्सीजिनेशन (ऑक्सीकरण) क्षमता भी होती है।
2. उपचयन (Reduction) – यह प्रतिक्रियाओं की एक शृंखला है जिसमें ग्लूकोज बनता है। इस चरण में प्रत्येक CO2 अणु के स्थिरण हेतु एटीपी के 2 अणुओं का उपयोग फॉस्फोरिलेशन के लिए तथा एनडीपीएच के दो अणुओं का उपयोग अपचयन हेतु होता है। पथ से ग्लूकोज के एक अणु को बनाने के लिए CO2 के 6 अणुओं के यौगिकीकरण तथा चक्करों की आवश्यकता होती है।
3. पुनरुद्भवन Regenertion – यदि चक्र को बिना बाधा के जारी रहना है तो CO2 ग्राही अणु RuBP का पुनरुद्भवन बहुत ही आवश्यक होता है। पुनरुद्भवन के चरण में RuBP गठन हेतु फॉस्फोरिलेशन के लिए एक एटीपी की आवश्यकता होती है।
कार्बन स्थिरीकरण के एकान्तरित चक्र (Alternative cycles of carbon fixation) –
– इसमें निम्नलिखित चक्र शामिल हैं–
(A) हेच तथा स्लेक चक्र या C4 – चक्र
(B) क्रेसुलेसियन अम्ल उपापचय या CAM चक्र
(A) हेच तथा स्लेक चक्र या C4 – चक्र (Hatch and slack cycle or C4 cycle) –
– हेच तथा स्लेक (1967) ने CO2 स्थिरीकरण के इस एकान्तरिक चक्र की खोज की जिसे हेच तथा स्लेक चक्र कहते हैं। इसे C4 – चक्र भी कहते हैं क्योंकि इस चक्र का प्रथम स्थाई यौगिक 4 C यौगिक ओक्जेलो एसीटिक अम्ल (OAA) होता है।
– C4 पादपों में पर्ण की पूलाच्छद कोशिकाओं में अभिकेन्द्रीय रूप से व्यवस्थित बड़े तथा एग्रेनल क्लोरोप्लॉस्ट (ग्रेना अनुपस्थित) रखते हैं। पर्णमध्योतक कोशिकाओं के क्लोरोप्लॉस्ट छोटे तथा उनमें ग्रेना पूर्ण विकसित होते हैं। पर्णमध्योतक कोशिकाएँ तथा पूलाच्छद कोशिकाएँ प्लाज्मोडेस्मेटा द्वारा जुड़ी होती है।
PEP कार्बोक्सिलेज एन्जाइम पर्णमध्योतक कोशिकाओं में पाया जाता है तथा RuBP कार्बोक्सिलेज पूलाच्छद में पाया जाता है। C4 – चक्र में निम्न चरण है–
(i) पर्ण मध्योतक कोशिकाओं में (In mesophyll cells) –
– C4 पादपों में CO2 सबसे पहले पर्णमध्योतक कोशिकाओं में फॉस्फोइनाल पायरुवेट (PEP) द्वारा ग्रहण की जाती है तथा चार कार्बन यौगिक ओक्जेलो एसीटिक अम्ल (प्राथमिक स्थायी यौगिक) बनता है।

OAA, 4C यौगिक मैलिक अम्ल में परिवर्तित हो जाता है। हाइड्रोजन NADPH + H+ द्वारा प्रदान की जाती है।

(II) पूलाच्छद में (In Bundle sheath) – मैलिक अम्ल अब पूलाच्छद कोशिकाओं में चला जाता है। जहाँ यह पायरुविक अम्ल में विकार्बोक्सिलीकृत हो जाता है तथा CO2 मुक्त होती है जो कि RUBP द्वारा ग्रहण की जाती है तथा केल्विन चक्र द्वारा हेक्सोज शर्कराओं के रूप में पुन:स्थिर की जाती है।

(III) पायरुविक अम्ल पर्णमध्योतक कोशिका में आता है तथा ATP की सहायता से फॉस्फोइनॉल पायरुवेट (PEP) में परिवर्तित हो जाता है।

– कुछ पादपों में एस्पार्टिक अम्ल पर्ण मध्योतक से पूलाच्छद में स्थानान्तरित होता है जहाँ यह मैलिक अम्ल में परिवर्तित हो जाता है तथा CO2 मुक्त होती है।
उदाहरण – पेनीकम मेक्सीमम।

C4 पादपों का महत्त्व (Significance of C4 plants) –
– हेच स्लेक चक्र राइबुलोज बाइफॉस्फेट के बजाय फॉस्फोइनॉल पायरुवेट के कार्बोक्सिलीकरण से प्रारम्भ होता है क्योंकि PEP कार्बोक्सिलेज RuBP कॉर्बोक्सिलेज की तुलना में CO2 के साथ अधिक बन्धुता दर्शाता है।
– C4 पादपों में पूलाच्छद का जाइलम के समीप स्थित होने से जल की कमी से होने वाला हानिकारक प्रभाव घट जाता है।
– C4 पादपों में उच्च प्रकाश संश्लेषी क्षमता के कारण तथा प्रतिकूल परिस्थितियों में जीवित रहने की क्षमता के कारण ये पादप खरपतवारों के रूप में विकसित होते हैं।
उदाहरण – चीनोपोडियम, एमरेन्थस, सालसोला।
– C4 पादप अधिकांशतः गर्म मरुस्थलों में पाए जाते हैं जहाँ उच्च प्रकाश तीव्रता तथा उच्च तापक्रम पाए जाते हैं। ये परिस्थितियाँ पादपों में प्रकाश श्वसन को प्रेरित करती हैं लेकिन C4 पादपों में प्रकाश श्वसन अनुपस्थित होता है।
– C4 चक्र RuDP के लिए अधिक CO2 प्रदान करता है। RUDP कार्बोक्सिलेज एन्जाइम CO2 के प्रति कम संवेदी होता है।
(B) क्रेसुलेसियन एसिड मेटाबॉलिज्म या CAM चक्र (Crassulacean Acid metabolism or CAM cycle)–

– ओलेरी, राऊनी तथा ब्लैक ने क्रेसुलेसी कुल के पादपों में सबसे पहले इसे प्रेक्षित किया था, इसलिए इसे CAM कहते हैं।
– CAM पादप, मुख्य रूप से गूद्देदार मरुद्भिद होते हैं।
उदाहरण – नागफणी, ब्रायोफिल्लम, अन्नानास, कलौंजी, सीडम, अगेव, एलोय, केसुला, युफोर्बिया स्पीशीज।
– इन पौधों में रंध्र रात्रि में खुलते हैं। इन रंध्रों को स्कोटोएक्टिव रंध्र कहते हैं।
– CAM पादपों की पत्तियों की पर्णमध्योतक कोशिकाओं में C3 तथा C4 चक्र के एन्जाइम्स होते हैं लेकिन केंज शारीरिकी अनुपस्थित होती है। इन पादपों में C3 चक्र दिन में जबकि C4 चक्र रात्रि में होता है।
– इस उपापचयी चक्र में शामिल हैं–
(i) अम्लीकरण (Acidification) – अंधेरे में रंध्र खुलने के कारण ग्लाइकोलिसिस की प्रक्रिया द्वारा संचित कार्बोहाइड्रेट्स पायरुवेट (PEP) में परिवर्तित हो जाते हैं। इस कारण CO2 पत्तियों में प्रवेश करती है तथा PEP, CO2 के साथ PEP कार्बोक्सिलेज एन्जाइम की उपस्थिति में क्रिया कर ओक्जेलो एसीटिक अम्ल (OAA) बनाते हैं। OAA, NADH+H+ की सहायता से एन्जाइम मैलिक डिहाइड्रोजिनेज की उपस्थिति में मैलिक अम्ल में अपचयित हो जाता है। यह मैलिक अम्ल रिक्तिका में संगृहीत हो जाता है।
(ii) विअम्लीकरण (Deacidification) – प्रकाश में मैलिक अम्ल विकार्बोक्सिलीकृत होकर पायरुविक अम्ल बनाता है तथा CO2 उत्पन्न होती है। इस प्रक्रिया को विअम्लीकरण कहते हैं। उपर्युक्त प्रक्रम में निर्मित CO2 सामान्य प्रकाश संश्लेषी प्रक्रिया में प्रयुक्त होकर कार्बोहाइड्रेट्स उत्पन्न करती है।
– इस प्रकार CAM पादपों में CO2 का स्थिरीकरण दिन तथा रात में दो बार होता है।
प्रकाश श्वसन (PHOTORESPIRATION) –
– C3 पादपों में प्रकाश की उपस्थिति में ऑक्सीजन द्वारा कार्बनिक यौगिकों का ऑक्सीकरण प्रकाश श्वसन कहलाता है।
– प्रकाश श्वसन का सब्स्ट्रेट ग्लाइकोलेट है, जो 2C यौगिक है इस कारण इसे C2 -चक्र या ग्लाइकोलेट चक्र भी कहते हैं।
– यह तीन भिन्न कोशिकांगों में होता है। viz- क्लोरोप्लॉस्ट, परासीसोम तथा माइटोकॉन्ड्रिया।
(1) क्लोरोप्लास्ट में (In chloroplast) – उच्च तापक्रम, उच्च प्रकाश तीव्रता, CO2 की कम सान्द्रता, O2 की उच्च सान्द्रता की उपस्थिति में, RUBISCO ऑक्साजिनेज के रूप में कार्य करता है। इसके कारण एक अणु 3 – PGA तथा एक अणु फॉस्फोग्लाइकोलेट बनता है।

2- फॉस्फोग्लाइकोलेट से अकार्बनिक फॉस्फेट पृथक् होकर ग्लाइकोलेट बनाता है जो परॉक्सीसोम में स्थानान्तरित हो जाता है।

(2) परॉक्सीसोम में (In peroxisome) – ग्लाइकोलेट ऑक्सीजन के साथ क्रिया कर ग्लाइऑक्सीलेट बनाता है जो ट्रांसएमीनेशन द्वारा ग्लाइसिन बनाता है।
(3) माइटोकॉन्ड्रिया में (In mitochondria)– अब ग्लाइसीन माइटोकॉन्ड्रिया में जाती है जहाँ ऑक्सीजन की उपस्थिति में दो अणु ग्लाइसीन क्रिया कर एक अणु सीरिन, CO2, NH3 बनाते हैं।
– सीरिन पुनः परॉक्सीसोम में चली जाती है जहाँ सीरिन का विएमिनीकरण (Deamination) होता है परिणामस्वरूप ग्लिसरेट बनता है जो क्लोरोप्लॉस्ट में पहुँच कर ATP की उपस्थिति में 3-PGA बनाता है। 3-PGA कार्बन स्वांगीकरण चक्र में प्रयुक्त होता है।
– प्रकाश संश्लेषण के दौरान स्थिर की गई 30 – 40% CO2 की प्रकाश श्वसन के दौरान हानि होती है।
प्रकाश संश्लेषण को प्रभावित करने वाले कारक (Factor affecting photosynthesis) –
– साक्स (1860) ने द्विगबिन्दु परिकल्पना (concept of cardinal point) प्रतिपादित की। इसके अनुसार किसी भी कार्यिकीय क्रिया को प्रभावित करने वाले कारकों के तीन मान होते हैं।
(a) न्यूनतम (Minimum) – कार्यिकी क्रिया शुरू नहीं होती यदि मान न्यूनतम से कम होता है।
(b) अनुकूलतम (Optimum) – अनुकूलतम मान पर, अभिक्रिया अपनी अधिकतम गति से अनिश्चित काल तक हो सकती है।
(c) अधिकतम (Maximum) – अधिकतम से अधिक मान होने पर क्रिया रुक जाती है।
सीमाकारी कारक का ब्लैकमैन का नियम (Blackman’s law of limiting factor) –
– इस नियम के अनुसार जब प्रक्रम अनेक कारकों द्वारा प्रभावित होता है तो प्रक्रम की दर सबसे कम परिमाण में उपलब्ध कारक द्वारा नियंत्रित होती है।
बाह्य कारक (External factors) –
(1) प्रकाश (Light) –
(A) प्रकाश तीव्रता (Light intensity) –
– प्रकाश संश्लेषण की दर विसरित प्रकाश की तुलना में तीव्र प्रकाश में अधिक होती है। प्रकाश संश्लेषण की दर प्रकाश की तीव्रता में वृद्धि के साथ बढ़ती है लेकिन उच्च प्रकाश तीव्रता में क्लोरोफिल का ऑक्सीकरण हो जाता है इस घटना को ‘सोलेराइजेशन’ कहते हैं तथा इसके कारण प्रकाश संश्लेषण की दर घट जाती है।
– दिन के समय श्वसन की तुलना में प्रकाश संश्लेषण की दर 10 गुना अधिक हो जाती है लेकिन शाम के समय एक बिन्दु पर प्रकाश संश्लेषण तथा श्वसन की दर समान होती है इसे प्रकाश संतुलन बिन्दु (light compensation point) कहते हैं। जो कि पादप के लिए अच्छा नहीं है।
(B) प्रकाश की गुणवत्ता (Quality of Light) –
– प्रकाश संश्लेषण की दर अधिकतम लाल प्रकाश में फिर नीले प्रकाश में होती है तथा यह हरे प्रकाश में न्यूनतम होती है। यह बहुवर्णी प्रकाश (polychromatic light) जैसे श्वेत प्रकाश (white light) में अधिकतम होती है।
(2) कार्बन डाईऑक्साइड (CO2) –
– CO2 की सामान्य सान्द्रता वायुमण्डल में 0.36% (360 ppm) होती है यदि CO2 की सान्द्रता बढ़ती है तो प्रकाश संश्लेषण की दर भी बढ़ती है लेकिन CO2 की उच्च सान्द्रता में यह घट जाती है।
– CO2 सन्तुलन बिन्दु (CO2 compensation point) – यह वह बिन्दु है जिस पर प्रकाश संश्लेषण में CO2 उपभोग की मात्रा तथा श्वसन में विमुक्त CO2 की मात्रा बराबर होती है। यह C3 पादपों के लिए 25-100ppm तथा C4 पादपों के लिए 0-10ppm होती है।
(3) तापक्रम (Temperature) –
– प्रकाश संश्लेषण के लिए अनुकूलतम तापक्रम 20o-35oC होता है लेकिन 40oC के ऊपर एंजाइम्स के विकृतीकरण के कारण प्रकाश संश्लेषण की दर में गिरावट आती है। प्रकाश संश्लेषण 0oC पर निरोधित हो जाता है लेकिन अपवाद स्वरूप लाइकेन्स तथा कोनीफर्स में 20 से 35oC पर भी प्रकाश संश्लेषण होता है। कुछ हरे-नीले शैवाल जैसे आसिलोटोरिया ब्रेविस उच्च तापक्रम (70oC) पर भी प्रकाश संश्लेषण करते हैं।
(4) ऑक्सीजन (O2) –
– प्रकाश श्वसन के कारण C3 पादपों में O2 की उच्च सान्द्रता की उपस्थिति में प्रकाश संश्लेषण की दर में गिरावट आती है, इसे वारबर्ग प्रभाव कहते हैं।
(5) जल (Water) –
– 1% अवशोषित जल प्रकाश संश्लेषण में प्रयुक्त होता है। जल की कमी के कारण रंध्र बंद हो जाते हैं तथा प्रकाश संश्लेषण की दर घट जाती है।
पादप वृद्धि
– एक जीवित वस्तु के लिए वृद्धि को सर्वाधिक आधारभूत एवं सुस्पष्ट विशिष्टता के रूप में जाना जाता है।
– वृद्धि को एक अवयव या अंग या इसके किसी भाग या यहाँ तक कि एक कोशिका के आधार में अनिवर्त्य (अनपलट) स्थाई बढ़त के रूप में परिभाषित किया जा सकता है।
– सामान्यत: वृद्धि उपापचयी प्रक्रियाओं (उपचय एवं अपचय दोनों) से जुड़ी होती है जो ऊर्जा के व्यय पर आधारित होती है। इसलिए एक पत्ती का विस्तार वृद्धि है।
– पादप वृद्धि अनूठे ढंग से होती है, क्योंकि पौधे जीवन भर असीमित वृद्धि की क्षमता को अर्जित किए होते हैं।
– इस क्षमता का कारण उनके शरीर में कुछ खास जगहों पर विभज्योतक (मेरिस्टेम) ऊतकों की उपस्थिति है।
– ऐसे विभज्योतक की कोशिकाओं में विभाजन एवं स्वशाश्वतता (निरंतरता) की क्षमता होती है, हालाँकि यह उत्पाद जल्द ही विभाजन की क्षमता खो देते हैं और ऐसी कोशिकाएँ जो विभाजन की क्षमता खो देती हैं, वे पादप शरीर की रचना करती हैं।
– इस प्रकार की वृद्धि जहाँ पर विभज्योतक की क्रियात्मकता से पौधे के शरीर में सदैव नई कोशिकाओं को जोड़ा जाता है, उसे वृद्धि का खुला स्वरूप कहा जाता है।
– वृद्धि की अवधि को मुख्यत: तीन चरणों में बाँटा गया है- विभज्योतकीय, दीर्घीकरण एवं परिपक्वता।
– विभज्योतकीय चरण में कोशिकाएँ मूल शिखाग्र तथा प्ररोह शिखाग्र दोनों में लगातार विभाजित होती हैं।
– इन क्षेत्रों की कोशिकाएँ जीवद्रव्य से भरपूर होती हैं और व्यापक संलक्ष्य केंद्रक को अधिकृत किए होती हैं।
– उनकी कोशिका भित्ति प्राथमिक, पतली तथा प्रचुर जीवद्रव्य तंतु संयोजन के साथ सेल्युलॉजिक होती है।
– विभज्योतक क्षेत्र के समीपस्थ (ठीक अगला, नोक से दूर) कोशिका दीर्घीकरण के चरण का प्रतिनिधित्व करता है। इस चरण में कोशिकाओं का बढ़ा हुआ रसधानी भवन, कोशिका विशालीकरण तथा नव कोशिका भित्ति निक्षेपण आदि विशिष्टताएँ हैं।
– पुन: शिखाग्र से आगे अर्थात् दीर्घीकरण के अधिक समीपस्थ अक्ष का वह भाग स्थित होता है जो कि परिपक्वता के चरण में जा रहा होता है।
– इस परिक्षेत्र में स्थित होने वाली कोशिकाएँ अपने अंतिम आकार को प्राप्त किए होती हैं तथा उनकी भित्ति की मोटाई एवं रसधानी चरम पर होती है।
– सर्वप्रथम Von Sachs ने वृद्धि व समय के बीच ग्राफ लिया था, जो S आकार का मिला, इसे ही वृद्धि वक्र या सिग्मॉयड वक्र या Grand period curve या S-वक्र कहा जाता है।
– कोशिकांग, कोशिका, अंग या जीवधारी के जीवन का ग्राफ ऐसा ही आता है।
– वृद्धि का ग्राफ, पौधों और जंतुओं दोनों में ऐसा ही आएगा, परंतु समय भिन्न-भिन्न हो सकता है। इसमें निम्नलिखित काल आते हैं-
1. Lag-period/phase- इस प्रारंभिक काल में वृद्धि धीमी होती है।
2. Log-period/phase- इसे exponential period भी कहते हैं, जिसमें वृद्धि तेज व अधिकतम होती है।
3. Stationary of steady phase- इसमें वृद्धि शांत होकर अंत में गिरती जाती है।
– वृद्धि की अवस्थाओं में लगने वाले समय (मुख्यत: Log phase) को वृद्धि का समग्र काल कहा जाता है।
पादप वृद्धि का मापन:-
– निम्नलिखित विधियों से किया जाता है-
1. प्रत्यक्ष प्रेक्षण (By direct observation)
2. क्षैतिज सूक्ष्मदर्शी द्वारा (By horizontal microscope)
3. क्रेस्कोग्राफ द्वारा (By Crescograph of Sir J.C. Bose) यह वृद्धि को 10,000 गुना बढ़ा कर दर्शाता है।
4. वृद्धिमापी द्वारा
(i) ARC auxanometer
(ii) Pfeffer or pully auxanometer स्थायी ग्राफ बनाता है।
(iii) Micrometer screw-auxanometer
वृद्धि को प्रभावित करने वाले कारक:-
प्रकाश:-
– प्रकाश, प्रकाश-संश्लेषण तथा प्ररोह एवं मूल की वृद्धि की दिशा को निर्धारित करने से संबंधित होता है।
– प्रकाश, वृद्धि की प्रारंभिक अवस्था एवं बीजांकुरण के दौरान आवश्यक नहीं होता।
– प्रकाश की अनुपस्थिति से पादपों में पाण्डुरता (etiolation) हो जाती है।
– प्रकाश, पौधों की अकारिकी का नियंत्रण करता है। इसे प्रकाश आकारिकीजन (photomorphogenesis) कहते हैं।
ताप:-
– वृद्धि के लिए अनुकूलतम ताप 20-35oC होता है।
– पादपों के पुष्पन पर न्यून ताप का प्रभाव वसंतीकरण कहलाता है।
जल:-
– कोशिका की स्फीति को बनाए रखना है जो कि वृद्धि के लिए आवश्यक है (स्फीत दाब या TP, वृद्धि के लिए महत्त्वपूर्ण होता है) वृद्धि के दौरान का मान शून्य नहीं होना चाहिए।
– जल जीवद्रव्य में एन्जाइम सक्रियता के लिए आवश्यक होता है।
ऑक्सीजन:-
– कोशिकीय श्वसन के लिए आवश्यक होती है।
खनिज पोषक तत्त्व:-
– ये सभी तत्त्व वृद्धि एवं उपापचय के लिए अनिवार्य होते हैं।
आधुनिक कारक:-
– Genotypic and Phenotypic
पादप वृद्धि नियामक:-
– पादप वृद्धि नियामक विविध रासायनिक संघटनों वाले साधारण तथा लघु अणु होते हैं।
– ये इंडोल सम्मिश्रण (इंडोल-3 एसिटिक अम्ल, आई.ए.ए.), ऐडनीन व्युत्पन्न, फरफ्युराइल ऐमिनो प्यूरीन काइनटिन केराटिनॉयड तथा वसा अम्लों के व्युत्पन्न (एब्सिसिक एसिड, ए.बी.ए.), टर्पीन (जिब्बरेलिक एसिड, जी.ए.) या गैसेस (एथीलिन C2H4) आदि हो सकते हैं।
– पादप वृद्धि नियामक को पादप वृद्धि, पादप हॉर्मोन तथा फाइटोहॉर्मोन के नाम से जाना जाता है।
– पादप वृद्धि नियामक (पी.जी.आर.) को व्यापक रूप से एक जीवित पौधे में उनकी कार्यशीलता के आधार पर दो समूहों में बाँटा जा सकता है।
– पी.जी.आर. का एक समूह वृद्धि उन्नयन क्रियाकलापों में लगा होता है जैसे– कि कोशिका विभाजन, कोशिका प्रसार, प्रतिमान संरचना, ट्रापिक (अनुवर्तनी) वृद्धि, पुष्पन, फलीकरण तथा बीज संरचना आदि। इन्हें पादप वृद्धि नियामक भी कहा जाता है। जैसे कि ऑक्सिंस, जिब्बरेलिन्स तथा साइटोकिनिंस।
– उनके समूह के दूसरे पी.जी.आर. तथा दबाव के प्रति पादपों की अनुक्रिया समूह के दूसरे पी.जी.आर. में महत्त्वपूर्ण भूमिका निभाते हैं।
– वे विभिन्न वृद्धि बाधक क्रियाकलापों जैसे प्रसुप्ति एवं विलगन में भी शामिल होते हैं।
– एब्सिसिक एसिड पी.जी.आर. इसी समूह का सदस्य है।
– गैसीय पी.जी.आर., एथीलिन किसी भी समूह के साथ बैठ जाता है लेकिन व्यापक तौर पर यह एक वृद्धि बाधक क्रियाकलापों में आता है।
– ऑक्सिन की खोज F.W. Went के द्वारा जई के अंकुर प्रांकुरचोल के शिखर से की गई है।
– ‘बैकेन’ (फूलिश सीडलिंग) धान के पौध (नवोद्भिद्) की बीमारी है जो रोगजनक कवक जिबेरेला फूजीकोराइ के द्वारा होती है।
– ई. कुरोसोवा (जापानी वैज्ञानिक) ने रोगरहित धान की पौध में रोग लक्षण को बताया, जब उन्हें कवक के जीवाणुहीन निस्यंदों (फिल्ट्रेट) के साथ उपचारित किया।
– सक्रिय तत्त्व की पहचान बाद में जिब्बरेलिक अम्ल के रूप में हुई।
– एच.एच. कजिन्स ने यह सुनिश्चित किया कि पके हुए संतरों से निकला हुआ एक वाष्पशील तत्त्व पास में रखे बिना पके हुए केलों को शीघ्रता से पकाता है। बाद में यह वाष्पशील तत्त्व एथीलिन के नाम से जाना गया।
ऑक्सिंस:-
– ग्रीक शब्द ऑक्सेन – बढ़ना। सर्वप्रथम मनुष्य के मूत्र से निकाला गया।
– शब्द ऑक्सिंस इनडोल-3 एसीटिक अम्ल (आई.ए.ए.) तथा अन्य प्राकृतिक एवं कृत्रिम यौगिक, जिसमें वृद्धि करने की क्षमता हो, के लिए प्रयोग किया जाता है।
– ये प्राय: तने एवं मल के बढ़ते हुए शिखर पर बनते हैं तथा वहाँ से क्रियाशीलता वाले भाग में जाता है।
– ऑक्सिंस; जैसे आई.ए.ए. एवं इनडोल ब्यूटेरिक अम्ल पौधे से निकाला गया है।
– एन.ए.ए. (नैफथेलिन एसेटिक अम्ल) तथा 2, 4 डी (2, 4 डाईक्लोरो फिनोक्सी एसेटिक अम्ल) कृत्रिम ऑक्सिंस हैं।
– ऑक्सिंस के उपयोग का एक विस्तृत दायरा है और ये बागवानी एवं खेती में प्रयोग किए गए हैं।
– ये तनों की कटिंग (कलमों) में जड़ फूटने (रूटिंग) में सहायता करती है जो पादप प्रवर्धन में व्यापकता से इस्तेमाल होती है। ऑक्सिंस पुष्पन को बढ़ा देती है, जैसे अनानास में।
– ये पौधों के पत्तों एवं फलों को शुरुआती अवस्था में गिरने से बचाते हैं तथा पुरानी एवं परिपक्व पत्तियों एवं फलों के विलगन को बढ़ावा देते हैं।
– उच्च पादपों में वृद्धि करती अग्रस्थ कलिका पार्श्व (कक्षस्थ) कलियों की वृद्धि को अवरोधित करते हैं, जिसे शिखाग्र प्रधान्यता (apical dominance) कहते हैं।
– ऑक्सिंस अनिषेकफलन को प्रेरित करता है जैसे कि टमाटर में।
– इन्हें व्यापक रूप से शाकनाशी के रूप में उपयोग किया जाता है।
– 2, 4-डी, व्यापक रूप से द्विबीजपत्री खरपतवारों का नाश कर देता है, लेकिन एकबीजपत्री परिपक्व पौधों को प्रभावित नहीं करता है।
– इसका उपयोग मालियों के द्वारा लॉन को तैयार करने में किया जाता है।
– इसके साथ ही ऑक्सिंस जाइलम विभेदन को नियंत्रित करने तथा कोशिका के विभाजन में मदद करता है।
जिब्बरेलिन्स:-
– जिब्बरेलिन्स एक अन्य प्रकार का प्रोत्साहक पी.जी.आर. है।
– कवकों और उच्च पादपों से।
– इन्हें जी ए1 (GA1), जी ए2 (GA2), जी ए3 (GA3) और इसी तरह से नामित किया जाता है हालाँकि जी ए3 वह जिब्बरेलिन्स है जिसकी सबसे पहले खोज की गई थी और अभी भी सभी से अधिक सघनता से अध्ययन किया जाने वाला स्वरूप है।
– सभी जी.ए.एस. (GAS) अम्लीय होते हैं। ये पौधों में एक व्यापक दायरे की कायिकीय अनुक्रिया देते हैं।
– ये अक्ष की लंबाई बढ़ाने की क्षमता रखते हैं, अत: अंगूर के डंठल की लंबाई बढ़ाने में प्रयोग किए जाते हैं।
– जिब्बरेलिन्स सेव जैसे फलों को लंबा बनाते हैं ताकि वे उचित रूप ले सकें।
– ये जरावस्था को भी रोकते हैं, ताकि फल पेड़ पर अधिक समय तक लगे रह सकें और बाजार में मिल सकें।
– जी ए3 (GA3) को आसव (शराब) उद्योग में माल्टिंग की गति बढ़ाने के लिए उपयोग किया जाता है।
– गन्ने के तने में कार्बोहाइड्रेट्स चीनी या शर्करा के रूप में एकत्र रहता है।
– गन्ने की खेती में जिब्बरेलिन्स छिड़कने पर तनों की लंबाई बढ़ती है।
– जी ए छिड़कने पर किशोर शंकुवृक्षों में परिपक्वता तीव्र गति से होती है अत: बीज जल्दी ही तैयार हो जाता है।
– जिब्बरेलिन्स चुकंदर, पत्तागोभी एवं अन्य रोजेटी स्वभाव वाले पादपों में वोल्टिंग (पुष्पन से पहले अंत:पर्व का दीर्घीकरण) को बढ़ा देता है।
साइटोकाइनिन:-
– साइटोकाइनिन अपना विशेष साइटोकिनेसिस (कोशिकाद्रव्य विभाजन) में डालता है और इसे काइनेटिन (एडेनिन का रूपांतरित रूप एक प्यूरीन) के रूप में आटोक्लेबड्र हेरिंग के शुक्राणु से खोजा गया था।
– काइनेटिन पौधों में प्राकृतिक रूप से नहीं पाया जाता है।
– प्राकृतिक साइटोकाइनिन उन क्षेत्रों में संश्लेषित होता है, जहाँ तीव्र कोशिका विभाजन सम्पन्न होता है, उदाहरण के लिए मूल शिखाग्र, विकासशील प्ररोह कलिकाएँ तथा तरुणफल आदि।
– यह नई पत्तियों में हरितलवक पार्श्व प्ररोह वृद्धि तथा अपस्थानिक प्ररोह संरचना में मदद करता है।
– साइटोकाइनिन शिखाग्र प्राधान्यता से छुटकारा दिलाता है।
– वे पोषकों के संचारण को बढ़ावा देते हैं जिससे पत्तियों की जरावस्था को देरी करने में मदद मिलती है।
एथीलिन:-
– एथीलिन एक साधारण गैसीय पी.जी.आर. है यह जरावस्था को प्राप्त होते ऊतकों तथा पकते हुए फलों के द्वारा भारी मात्रा में संश्लेषित की जाती है।
– एथीलिन पौधों की अनुप्रस्थ (क्षैतिज) वृद्धि, अक्षों में फुलाव एवं द्विबीजी निवेद्भिदों में अंकुश संरचना को प्रभावित करती है।
– एथीलिन जरावस्था एवं विलगन को मुख्यत: पत्तियों एवं फूलों में बढ़ाती है।
– यह फलों को पकाने में बहुत प्रभावी है।
– फलों के पकने के दौरान यह श्वसन की गति की वृद्धि करता है।
– श्वसन वृद्धि में गति की इस बढ़त को क्लाइमैक्टिक श्वसन कहते हैं।
– एथीलिन बीज तथा कलिका प्रसुप्ति को तोड़ती है, मूँगफली के बीज में अंकुरण को शुरू करती है तथा आलू के कंदों को अंकुरित करती है।
– एथीलिन गहरे पानी के धान के पौधों में पर्णवृंत को तीव्र दीर्घीकरण के लिए प्रोत्साहित करता है।
– यह पत्तियों तथा प्ररोह के ऊपरी भाग को पानी से ऊपर रखने में मदद करता है।
– इसके साथ ही एथीलिन मूल वृद्धि तथा मूल रोमों को प्रोत्साहित करती है, अत: पौधे को अधिक अवशोषण क्षेत्र प्रदान करने में मदद करती है।
– एथीलिन अन्नानास को फूलने तथा फल समकालिकता में सहायता करता है। इसके साथ ही आम को पुष्पित होने में प्रेरित करता है।
– सर्वाधिक व्यापक तौर पर इस्तेमाल होने वाला यौगिक एथिफॉन है।
– एथिफॉन जलीय घोल में आसानी से अवशोषित तथा पौधे के अंतर्गत संचारित होता है तथा धीरे-धीरे एथीलिन मुक्त करता है।
– एथिफॉन टमाटर एवं सेव के फलों के पकाने की गति को बढ़ाता है तथा फूलों एवं फलों में विलगन को तीव्रता प्रदान करता है।
एब्सिसिक:-
– सर्वप्रथम निरोधकों का संकेत Osborne द्वारा दिया गया।
– Bennet-Clark व Kefford (1953) ने भी प्रसुप्त आलू के कंद से सर्वप्रथम वृद्धि संदमक प्राप्त किया जिसे उन्होंने 𝛽-inhibitor कहा था।
– Addicott व Okhuma (1963) ने कपास के परिपक्व फलों के रस से एक वृद्धि निरोधक प्राप्त किया, जो विलगन प्रेरित करता था व “Abscisin-II” कहा गया।
– Waring & Robinson ने भी Betula की पुरानी पत्तियों से एक growth-inhibitor पृथक् किया जो सुप्तावस्था प्रेरित करता था और इसे ‘dormin’ कहा गया।
– रासायनिक विश्लेषण से पता चलता है कि abscisin-II, 𝛽-inhibitor व dormin एक ही पदार्थ Abscisic acid (ABA) है।
– ABA को (Stress hormone) तनाव हॉर्मोन भी कहा जाता है, क्योंकि यह विपरीत परिस्थितियों में बनकर पौधों की रक्षा भी करता है।
– जल की कमी से रंध्र बंद करना। गर्मियों में वनों में विलगन करना आदि।
– (anti transpirant hormone है) ABA पौधे की विभिन्न तनावपूर्ण परिस्थितियों से रक्षा करता है।
– पादपों में सामान्य वृद्धि निरोधक ABA ही होता है।
ABA के कार्यिकी प्रभाव व महत्त्व:-
विलगन (Abscission) (ABA) का प्रमुख कार्यबद्ध का प्रेरण:-
– ABA आयु तथा विलगन का प्रेरक होता है।
– यह Cellulase व Penctinase genes को सक्रिय करके पुराने अंगों को मातृ पौधे से अलग करवाता है।
कलिका (Bud) व बीज (Seed) प्रसुप्ति dormancy का प्रेरण:-
– ABA seeds व buds की प्रसुप्ति को नियंत्रित करता है। (anti GA)/ABA बीजों के परिपक्वन में महत्त्वपूर्ण भूमिका निभाता है व बीजों में प्रसुप्ति करता है।
जीर्णता:-
– ABA पौधों में जीर्णता को त्वरित करता है।
कोशिका विभाजन व दीर्घीकरण का विरोध:-
– Cytokinin, Auxin (anti CK & anti Auxin) के विपरीत कार्य करता है।
रंध्रों को बंद करना:-
– ABA द्वार कोशिकाओं में Carotenes के ऑक्सीकरण से भी बनता है व रंध्रों को बंद करता है।
– शुष्क कृषि में उपयोग किया जा सकता है।
LDP के पुष्पीकरण में विलम्ब:-
– GA (anti GA) के विपरीत कार्य।
बीज अंकुरण का निरोधी:-
– ABA, 𝛼-amylase आदि के संश्लेषण का विरोधी है, अत: बीजों के अंकुरण को रोकता है।
– जड़ों में गुरुत्वानुवर्तन का प्रेरण।
– आलू में कंद निर्माण का प्रेरण।
– Duckweed (Lemna) में वृद्धि का विरोध करता है।
One thought on “पादप कार्यिकी”